| Paradoxical Sleep - A Study of its Nature and Mechanisms |
| Michel Jouvet Progress In Brain Research Vol. 18 Sleep Mechanisms 1965 |
Introduction
It has recently been discovered that during behavioural sleep there periodically occurs a state characterized by fast cortical activity similar to that of the waking state, accompanied by a complete disappearance of muscular tonus and of rapid eye movements (Dement, 1958; M. Jouvet et al., 1959). This gives rise to the following problem: Should we consider "classic" sleep, with its slow cortical waves (slow sleep), and paradoxical sleep (P.S.) to be the expression of a single hypnogenic mechanism? On the basis of this hypothesis, P.S. would represent no more than the overstatement of an active hypnic process, since in adults P.S. almost always follows slow sleep and is the deepest state of sleep ( M. Jouvet et al., 1959; Hubel, 1960; Rossi et al., 1961).
Or, on the other hand, should we consider P.S. to be a specific state of central nervous activity which differs qualitatively from slow sleep ? Such a dualistic hypothesis must be based on the experimental differentiation of P.S. and slow sleep. To this end, we shall present tile EEG findings and behavioural, phylogenetic, ontogenetic, functional and structural criteria, which represent a number of concordant arguments in favour of the duality of the states of sleep and of the autonomy, at least relative, of P.S. with respect to slow sleep. In the second part we shall attempt to delimit certain mechanisms regulating the appearance of P.S.
Evidence of the duality of the states of sleep
(a) EEG and behavioural findings
It is believed at the present time that slow sleep is characterized (in the chronic adult cat) by two successive EEG stages. The first is made up of cortical spindles of 15-18 c/s, the second of slow high-voltage waves of 2-4 c/s which invade the cortex and subcortical structures. These two stages are accompanied by tonic nuchal activity in the EMG and almost complete absence of eye movements. The appearance of spindles requires the integrity of the thalamus (Naquet et al. 1964), while that of slow cortical and subcortical waves necessitates the neocortex (M. Jouvet, 1962a). The mechanisms of the appareance of slow sleep have been discussed elsewhere (M. Jouvet, 1962a; Moruzzi, 1960).
Paradoxical sleep differs entirely from slow sleep in EEG, behavioural, tonic and phasic aspects.
(i) Tonic aspects (Fig. 1)
(1) EEG. ln normal adult cats not deprived of sleep, P.S. always occurs after a phase of slow sleep. Its average duration is 6 min, but periods of 15-20 min are often recorded. It represents an average proportion of 20-25% of behavioural sleep (i.e. approximately 15% of the total time). lt is characterized by fast, low-voltage neocortical, diencephalic and mesencephalic activity (20-30 c/s), similar to that of cortical activation which regularly accompanies a state of intense alertness or attention. But certain local electrical peculiarities make it possible to draw a formal distinction between the electrical cerebral activity of P.S. and that of behavioural alertness. The appearance of continuous 6-activity in the dorsal and ventral hippocampus is highly characteristic: it is more regular, more rapid (5-7 c/s) and, above all, of a more extensive topography than that observed during intense alertness (4-4.5 c/s) in the dorsal hippocampus�the presence of a 6-rhythm in the ventral hippocampus occurring only exceptionally in the waking state. A 6-rhythm has also been registered in the peri-aqueductal grey matter, the anterior portion of the pons and the limbic midbrain area (M. Jouvet, 1962a).
(2) Behavioural criteria. In contrast to slow sleep, which is not clearly defined behaviourally, the beginning and end of P.S. can be located to within a few seconds on the basis of behavioural criteria alone. The complete extinction of muscular activity, especially of the neck, is the most striking sign of the inhibition of muscular tonus that characterizes P.S. (M. Jouvet et al., 1959). Before, or some seconds after the cortical activity of P.S. begins, the lack of activity in the EMG is accompanied by a sudden dropping of the animal's head if it has been in an unsupported position during slow sleep. The end of P.S. is marked by a renewal, usually sudden, of considerable activity in the EMG, the awakening of the animal or transition back to the state of slow sleep.
(ii) Phasic aspects
Among the phasic phenomena characteristic of P.S. the eye movements are of such great importance that we shall consider them separately. But they are not in fact an isolated phenomenon and P.S. is punctuated in a strange and disordered manner by sudden movements of the ears, whiskers, limbs (flexion) and tail, and sometimes veritable clonic jerks of the muscles of the back (Fig. 2). These phasic phenomena are particularly developed in the young cat after birth and are characteristically increased after long privation of P.S.
Phasic phenomena of the oculomotor system. One of the most remarkable characteristics of P.S. is the appearance of rapid eye movements, accompanied by phasic ponto-geniculo-visual electrical activity.
Rapid eye movements (Fig. 3). Rapid eye movements occur from the beginning of cortical activation. With a frequency of 60-70 per min, they differ in speed, distribution, and pattern from ocular movements of observation during the waking state (Jeannerod and Mouret, 1963). They may be isolated or occur in short bursts of less than 5 movements (as during observation), but most characteristic are bursts of a greater number of movements, up to 50 without a pause. The ratio between the number of movements during the bursts and the total number of movements remains constant for each animal (50%). During P.S., myosis is at a maximum most of the time while the nictitating membranes are relaxed. Nevertheless, sudden mydriasis with retraction of the nictitating membranes can on occasions accompany the bursts of eye movements (Berlucchi et al., 1964). Analysis of the structures responsible for the appearance of isolated movements and bursts gave the following results (Jeannerod et al., 1965): pontile cats in which the superior colliculi were destroyed showed only isolated lateral and external movements (dependent on N. VI). In the mesencephalic cat with superior colliculi intact the larger bursts persisted. In contrast, coagulation of a zone in the superior colliculus and the mesencephalic tegmentum in intact animals suppressed these bursts. The latter were, on the other hand, very much enhanced in the totally decorticated animal. The role of the cortex is not unequivocal, for ablation of the visual cortex will decrease the number of bursts and isolated movements, sometimes to a considerable degree, whilst frontal decortication increases the number of bursts.
These observations can be summarized as follows: the rapid eye movements in P.S. are not identical with those of the waking state and persist, for example in decorticated, pontile cats, when eye movements in the waking state are completely impossible. They are also present during P.S. in kittens immediately after birth and still blind (Valatx et al., 1964). The mechanisms responsible for the rapid eye movements of P.S. must thus be different from those regulating the eyes during observation. The results suggest that these eye movements are initiated in the pons and are rendered more complex in the superior colliculi and the midbrain, whilst the process of 'cortical integration' (facilitating visual cortex and inhibiting frontal cortex) would act on this latter region.
Phasic ponto-geniculo-occipItal. activity (Fig. 4). Owing to the difficulties encountered and the slow development of a broad and systematic study of the cortical and subcortical structures under chronic conditions several years were necessary before a common link was found between the 'spontaneous' potentials observed during eye movements in P.S. First observed in the pontine reticular formation, monophasic peaks of 200 300 microV of 100 msec duration, often appearing in groups of five or six (whence their appearance as pseudo-spindles) (M. Jouvet et al., 1959), were then observed in the lateral geniculate nucleus (Mikiten et al., 1961), the occipItal. cortex (Mouret et al., 1963), the superior colliculus and oculomotor nucleus (Brooks and Bizzi, 1963; Michel et al., 1964a), and the pulvinar and parietal cortex (Hobson, 1964). The pontine and geniculate spikes are the earliest signs of incipient P.S. They can in fact precede rapid cortical activity and extinction of the nuchal EMG by several seconds. More rarely, this phasic activity can occur in fleeting bursts during slow sleep without the appearance of P.S.
Lesions of the nucleus reticularis pontis caudalis or in front of the pons, in the dorsal or central part of the brain stem, may suppress the appearance of geniculate and visual spikes during P.S. (Hobson, 1964). On the other hand, monophasic spikes persist in the pontine reticular formation (PRF) in the pontile animal during P.S. lt is thus probable that an ascending ponto-geniculo-occipItal. system, of which the topography has still to be clarified, reacts phasically during the rapid eye movements.
But the relationship between this phasic activity and the eye movements is not a simple one. Neither darkness, retinal coagulation, nor even total ablation of the eyeballs and the extrinsic muscles of the eye (Michel et al., 1964a) suppresses the pontovisual peaks, which therefore cannot be regarded as a possible feedback of retinal (on and off effect) or extrinsic muscular origin (Fig. 4). Moreover, this phasic activity precedes the eye movements by some 30-90 sec at the beginning of P.S., and the movement can occur without demonstrable spike activity, but in the majority of cases there is a relation in time between the monophasic ponto-geniculo-occipItal. spike and the activity of the extrinsic muscles of the eye. This activity appears above all as rapid phasic bursts, whilst in the waking state a tonic element occurs (Michel et al., 1964b).
It would be premature to try to correlate this phasic activity and the rapid eye movements at the present time, and it is sufficient to note the essential difference between these phenomena and those occurring during the eye movements of observation (when such phasic activity is not recorded) and especially during slow sleep, during which phasic phenomena occur neither in the motor effectors nor in the EEG.
These findings thus enable us to draw a clear distinction between P.S. and slow sleep on the basis of their EEG and tonic and phasic behavioural aspects. By these criteria P.S. appears as distinct from slow sleep as the latter is from the waking state.
But we cannot affirm on the basis of EEG methods and polygraphy alone that slow sleep and P.S. are the result of different mechanisms and structures. In order, therefore, to obtain more evidence in favour of the dualist concept for the two states of sleep ( M. Jouvet et al., 1959) we studied the possibility of differentiating them, either in the course of their phylogenic or ontogenic evolution, by selective deprivation, or by central nervous lesions.
Evidence of the duality of the states of sleep
(b) Phylogenetic findings
Polygraphic studies of sleep in vertebrates (Klein, 1963; Klein et al., 1964; Hermann et al., 1964) provide comparative physiological evidence on the basis of which the phylogenetic evolution of the two states of sleep can be differentiated. Slow sleep has in fact been demonstrated in all the mammals studied so far (see bibliography, Jouvet and Jouvet, 1964). It is also very readily recognizable in birds and reptiles by the presence of slow waves in the hyperstriatum of hens or pigeons and the archipallium of tortoises in association with immobility, closing the eyes, slowing down the respiratory and cardiac function, and the preservation of a certain muscular tonic activity in the neck. In contrast, during behavioural sleep in the tortoise no periods of rapid electrical archipallial activity associated with eye movements can be demonstrated. Paradoxical sleep thus apparently does not occur in chelonians. In birds (pigeon, hen, chick), on the other hand, very short periods of P.S. occur, lasting from 6-15 sec (Figs. 5 and 6).
They are characterized by the appearance, after a phase of slow sleep, of rapid activity in the hyperstriatum, rapid eye movements, an appreciable - but not complete - reduction in nuchal EMG activity, considerable bradycardia and postural relaxation objectified by drooping of the wings. These phases of rudimentary P.S. thus constitute only 0.15 - 0.2% of the behavioural sleep. In mammals, on the other hand, P.S. is much more developed and accounts for 6-30% of the behavioural sleep of adults, depending on the species (Fig. 25).
Evidence of the duality of the states of sleep
(c) Ontogenetic findings
Investigations into the ontogenetic evolution of slow and paradoxical sleep have been described in detail elsewhere (Valatx et al., 1964; D. Jouvet et al., 1961). The results show three categories of facts supporting the theory of distinct mechanisms for P.S. and slow sleep:
(i) During the first days after the birth of kittens, P.S., known as 'sleep with jerks' (Valatx et al., 1964) or 'agitated sleep' (Cadilhac et al., 1961), accounts for almost all (80-90%) the behavioural sleep. lt is characterized by behavioural phenomena (global muscular jerks, rapid eye movements, extinction of nuchal muscular tonus) in which the phasic aspect is far more pronounced than in adult animals. The average duration of P.S. at one week of age is similar to that of the adult cat, while its frequency is higher (Fig. 7). Thus, the mechanism responsible for P.S. is present from the very start of life, whereas the state of slow sleep is hardly recognizable at this stage.
(ii) P.S. can appear immediately after behavioural awakening during the first days of life, without any transitional phase of slow sleep.
(iii) During maturation the two states of sleep develop differently: the periods of P.S., of which the average duration remains constant, become less and less frequent, and constitute only 25-30% of behavioural sleep in the adult; the duration of slow sleep, on the other hand, increases from 10% at birth to 70% of behavioural sleep in the adult (Fig. 8).
Evidence of the duality of the states of sleep
(d) Functional findings
A technique of instrumental and selective deprivation of P.S. (D. Jouvet et al., 1964) was employed in order to dissociate the two states composing behavioural sleep in the adult cat. The animal is placed on a small support floating on water. It can stand or crouch, but the small surface of the support prevents it from Iying down completely and relaxing its muscular tonus without falling into the water. The EEG and EMG activity is recorded continuously or integrated by means of an Oneirograph (M. Jouvet, 1962b). The animal's behaviour is further recorded with the aid of photographs.
Four cats were subjected to successive periods of privation of 10, 24, 36, 48, 72 and 96 h, and 9 and 17 days. A minimum of a week was left between any two sessions to permit complete recuperation.
Results of deprivation:
(i) At the beginning of deprivation, behavioural and EEG arousal was slightly increased (40-60%) as a result of agitation, but phases of slow sleep reappeared within about twelve h. Spindles and even slow waves appeared in the cortex and subcortical structures, while the neck of the animal flexed. These phases of slow sleep were always followed by a sudden arousal caused by loss of balance as the neck bent more and more. Behavioural or EEG P.S. is thus impossible. Whilst deprivation of P.S. is absolute, deprivation of slow sleep is minimal (10-20% depending on the animal). Even during the longest of these periods of deprivation we never observed hallucinatory patterns like those occurring after lesions of the pontine reticular formation which suppress P.S. (M. Jouvet, 1962a). There was a marked increase in the pulse rate.
(ii) Recuperative phases were identical in all the animals. On leaving the tank, even after deprivation for as long as 17 days, the animals always indulged in a stereotyped act of grooming for 30 min to 1 h, after which they fell into a very deep sleep. On awakening their behaviour was reminiscent of asthenia. They were unable to jump onto a chair to obtain food and fell heavily to the ground. The first 6 h of recuperative sleep are represented in Fig. 9: after a deprivation of 72 h a plateau of 60% P.S. (in terms of behavioural sleep) was reached, and was not exceeded even when deprivation lasted for 17 days. This high percentage of P.S. is due to a slight increase in its average duration (8 min against 6 min 20 sec in controls) and especially to the shorter intervals between the phases of P.S. During the first episodes of recuperative P.S. the twitching of the animal's body, paws, tail, and whiskers, was so intense that the animal occasionally presented a picture of epileptic seizures. At these times the arousal threshold is very high and nociceptive stimuli are necessary to awaken the animal, while acoustic stimuli have no effect. The explosive return of P.S. after deprivation thus suggests a phenomenon of rebound and elective recuperation. The relative increase in P.S. during recuperative sleep is proportional to the duration of the deprivation period. It was 20, 60 and 200 h for periods of 2, 5 and 17 days of deprivation respectively. Another important finding was that after deprivation of more than 3 days P.S. can immediately follow the waking state without spindles or slow waves to characterize a transitory phase of slow sleep, as would be the case in the normal animal. Thus total deprivation of P.S. produces a large, lasting and selective increase in P.S. during recuperation. This also speaks in favour of the existence of specific mechanisms for P.S. distinct from those for slow sleep.
Evidence of the duality of the states of sleep
(e) Structural findings
(i) Lesions of tlte pontine reticular formation
We have reported elsewhere the results of coagulation of the pontine reticular formation at the level of the dorsal and lateral part of the nucleus reticularis pontis oralis et caudalis (M. Jouvet, 1962a). There is no extinction of the nuchal EMG in these animals nor periods of rapid activity during behavioural sleep. On the other hand, typical EEG patterns of wakefulness and slow sleep persist. When deprived of P.S. these animals show periodical behavioural disturbances resembling hallucinations that occur most commonly after a phase of slow sleep. The percentage of slow sleep (60% of the total time) was usually normal in these animals (Fig. 10). These results speak in favour of a duality of the nervous structures responsible for triggering off the two states of sleep. Study of the waking-sleeping rhythm of chronic pontile cats strengthens this theory.
(ii) Sleep in the chronic pontile cat
The experimental evidence by which the periodic phases of atony in chronic mesencephalic or pontile animals can be identified with the phase of fast cortical activity in intact animals during sleep, has already been reported (M. Jouvet, 1961, 1962a). It is on these results and the results of coagulation of the pontine reticular formation that the concept of rhombencephalic sleep is based. But the periodic atonic state of pontile animals cannot be unreservedly identified with P.S. (Moruzzi, 1964).
Our initial results had been obtained in animals which did not survive more than 10 days, and we therefore felt it necessary to obtain new data from animals with a much longer period of survival. Better knowledge of their requirements enabled us to study some 20 posterior mesencephalic or pontile animals for over 2 months. For such periods it is much easier to observe the different states of wakefulness. In a number of animals it was further possible to study more extensively the electrical activity of the brain stem.
Our technique was very similar to that developed by Bard and Macht (1958). After total section of the brain stem in front of the tentorium with a leukotomy knife, the cerebral hemispheres and the thalamus are removed by aspiration. A hypothalamic island is left in situ, the dorsal surface of which is flush with the Horsley-Clark plane zero. The brain stem section between the pons and the remaining hypothalamic island is then completed by aspiration and an acrylic strip 1-2 mm thick and 15 mm wide, fixed dorsally to the tentorium cerebelli, is wedged between the brain stem section surface and the remaining hypothalamic island in such a manner as to prevent any neurocrine connections between the hypothalamus and the brain stem (Fig. 11). These poikilothermic preparations are then placed in an incubator and observed through a perspex window. A thermo-electric probe in the rectum of the animal conveys its temperature to a regulator (Fig. 12, B). The animal's temperature is thus maintained constant at any desired level by altering the heating element of the incubator. The urine is collected (K). The EEG activity of the brain stem (pons and medulla oblongata) and the EMG activity of the neck, are registered by permanently implanted electrodes and transmitted by cable I to the amplifiers (C) of the EEG apparatus (D). After amplification the muscular activity is integrated in an Oneirograph (F). During P.S. the absence of EMG activity is conveyed to a system of relays (G) so that either the EEG motor (E) is automatically set in operation to record P.S. during the night, or the periodicity of P.S. is registered by a signal on a slow-running apparatus (H). The sleeping-waking rhythm of the animals can thus be continuously recorded. More than 10 000 periods of P.S. have thus been observed in 20 chronic pontile cats (Fig. 13).
(iii) Results
Immediately following the postoperative phase (after 12-36 h) the animals alternately presented wakefulness and P.S. without slow sleep intervening.
Wakefulness is characterized by muscular hypertonia with regular respiration - a complete absence of movements over the first few days. This state is thus difficult to identify since behaviourally it can resemble calm sleep during the first week. After about 10 days, however, this state can be regarded as a state of wakefulness. Theanimal then responds to acoustic stimuli of high intensity by turning its head towards the stimulus. Furthermore, as observed by Bard and Macht (1958), the animals are able to remain in a crouching position, with the head raised, supporting themselves on their forelegs (Sphinx position). In this state the EEG of the brain stem shows continuous, fast, microvoltage activity which is not affected by any sensory stimuli.
The waking state is periodically and regularly interrupted (Fig. 13) by a state of which all the characteristics are similar with those of P.S. in intact cats. This state consists of complete muscular atony, during which the head falls abruptly and eye movements appear. These movements are lateral (i.e. dependent on N. VI) external, rapid, returning slowly to the median line with a frequency of 20-30 per min. Their oculographic appearance is monotonous, in contrast to those of intact cats where several types of bursts of varying complexity can be recognized. During this state respiration becomes more rapid and irregular, while the heart rate increases in most cases. At the same time a characteristic electrical activity appears in the pons (Fig. 14): either isolated monophasic spikes or groups of 'pseudo spindles', 3-5 per sec, repeated at a frequency of 10-15 per min, sometimes rising in amplitude, seldom rising and falling. In the majority of cases these spikes accompany the rapid eye movements, but not infrequently, especially in the first few days, there are isolated spikes without eye movements. The topographical distribution of these elements, and their electrical appearance, is similar to that in the pontine reticular formation during P.S. (M. Jouvet, 1962a; Brooks and Bizzi, 1963).
(1) P.S. occurs immediately after the waking state. It has, in fact never been possible to demonstrate EEG or behavioural criteria of slow sleep in pontile animals.
Whereas in intact cats the electrodes located in the pontine reticular formation regularly receive spindle activity and/or slow waves during the slow sleep preceding the appearance of P.S., no slow activity or spindles have ever been recorded during the minutes preceding P.S. in pontile animals. Furthermore, no behavioural criteria have been observed for a stage intermediate between wakefulness and P.S. The state of the pupils and the nictitating membranes remains constant in these animals. In certain cases - mesencephalic animals with intact oculomotor nucleus - a tonic ocular sleep syndrome (in which the eyeballs rock in and down) can appear. This phenomenon immediately precedes, by a few seconds, the rapid eye movements of P.S. It has never been observed alone during long periods without P.S. Furthermore in 90% of such animals muscular activity remains constant during the waking phases and begins to diminish only 30-40 sec before the inception of P.S., when the first monophasic spikes appear in the pons. In the normal animal, on the other hand, the nuchal EMG activity diminishes considerably in about 60% of cases during the phase of slow sleep preceding P.S. Thus, the muscular criterion of slow sleep (which even in intact animals is not absolute and seems to depend on the animal's posture and thus often on the environmental temperature) does not show the hypotonic phases which would precede P.S. and would provide evidence of an intermediary behavioural sleep between wakefulness and P.S.
(2) Periodicity of P.S. The mean duration of P.S. in pontile and intact cats is the same (6 min and 6 min 20 sec respectively). Thus the two phenomena, which are identical from the point of view of subcortical electrical activity and of behaviour, are also identical from the point of view of time and can be entirely equated one with the other. The only difference is found in their periodicity. In pontile cats it is very regular and P.S. accounts for 10% of the total time (there is no distinction between day and night). In normal animals, on the other hand, the innumerable environmental influences (to which the pontile cat is almost totally insensitive) make it difficult to establish a regular periodicity of P.S. Furthermore, P.S. occurs only after periods of slow sleep when it accounts for approximately 25% of behavioural sleep, or 15% of the total time (since intact cats slept for an average of 70% of the time under the conditions of our trial, i.e. in sound-proof cages to which they have become 'accustomed').
Summarizing, no EEG or behavioural phenomenon indicates the existence of slow sleep in pontile animals. On the other hand, the periods of muscular atonia occurring in such animals immediately after awakening can be completely equated with the P.S. of intact animals. Their average duration is the same as the latter, while the percentage of the total time they represent is slightly lower. P.S. thus appears to be absolutely independent of slow sleep.
After having discussed in this first section the autonomy of P.S. with respect to slow sleep we shall pass in the second to some experimental results which make it possible to define some of the mechanisms at work. Most of these results have been obtained in pontile animals, in which P.S. appears as a veritable 'biological clock', the periodicity of which is subject to fewer factors than in the intact animal.
The possibility of triggering off P.S. as a reflex suggests that peripheral mechanisms may be involved, but not exclusively, as the results of different deafferentations have shown.
Experiments with automatic deprivation of P.S. in pontile animals suggest the existence of an active mechanism in the lower brain stem.
Finally, the effects of temperature and certain drugs, and the relationship of P.S. with the internal milieu permit the hypothesis of a periodic function in which the neuroglia is perhaps implicated.
Mechanisms of paradoxical sleep
(a) Producing P.S. as a reflex
The immediate triggering of P.S. has been obtained during slow sleep in intact animals by stimulation of the pontile reticular formation (M. Jouvet, 1961), mid-brain reticular formation (Rossi et al.,1961), and hippocampus (Cadilhac et al., 1961). The same phenomenon has been seen following stimulation of the pontine reticular formation in pontile and mesencephalic animals and the existence of refractory phases after phases of spontaneous or provoked P.S. was also noted (M. Jouvet, 1961). Furthermore, periods of slow sleep can be induced by low-frequency stimulation of the cutaneous or muscular nerves during wakefulness (Pompeiano and Swett, 1962a,b), whilst the same stimulation during slow sleep in intact cats only exceptionally produces P.S. (Pompeiano, 1964).
In the pontile animal, on the other hand, the P.S. reflex can regularly be obtained by proprioceptive and nociceptive stimulation. This phenomenon occurs only when certain conditions are fulfilled.
Anatomical conditions: Reflex P.S. occurs only when section of the brain stem is placed behind the origin of III. If the section is at or in front of this point in the mesencephalic animal, nociceptive and proprioceptive stimuli increase tonus and rigidity. Fig. 15 shows the most posterior (A) section to prevent the P.S. reflex;
(B) and (C) indicate the anterior and posterior limits of sections compatible with the appearance of the P.S. reflex and (D) the level of the section in retropontile animals in which no P.S. reflex could be obtained. These sections thus delimit two zones: the first, anterior to (A), at the level of the mesencephalic tegmentum, the integrity of which prevents the appearance of the P.S. reflex; these cond, between (C) and (D), at the level of the pons, which is necessary for the reflex and which encompasses the posterior part of the nucleus reticularis pontis oralis and the anterior part of the N.R.P. caudalis (based on the coordinates of the atlas of Snyder and Niemer, 1961). (It should be noted that the coordinates of the reticular formation nuclei of the pons vary from atlas to atlas. Thus, the anterior limits of the nucleus reticularis pontis caudalis are situated at P2 according to Snyder and Niemer, (1961) and at P5 according to Reinoso-Suarez (1961).)
Stimuli: P.S. can be triggered off as a reflex by the following stimuli: opening the mouth, introducing a tube into the esophagus with or without fluid, pinching the ear or fore or hind paws, flexion, extension, passive rotation of the head, passive flexion or hyperextension of the limbs. Cutaneous stimuli (stroking the back, face, stomach, limbs), and auditive stimuli, on the other hand, had no effect. Nociceptive stimuli almost always produce an immediate extension reaction of very short duration (a few seconds) and very brief apnea, and it is possible that the common denominator of all these reactions is the involvement of proprioceptive afferents.
The P.S. obtained by these stimuli appears either immediately (after 1-3 sec) or after a delay of 20-30 sec. It is accompanied by the same EEG, autonomic and behavioural (eye movements) phenomena as spontaneous P.S., and the duration of both is identical. A 'refractory period' of 10-20 min follows spontaneous or evoked P.S., during which the same stimuli either are totally ineffective or produce phasic suppression or, more rarely, tonic suppression of the EMG (cataplexy), which can last for 1 or 2 min. But then no monophasic spikes occur in the pons and there are no eye movements (Fig. 16). Such cataplexic periods cannot therefore be identified with P.S. The duration of the refractory phases following nociceptive and proprioceptive stimulation was identical. It was shorter than the mean interval occurring between two phases of spontaneous P.S. Finally intravenous injection of 1-5 microg/kg adrenalin never caused a P.S. reflex in these animals.
Mechanisms of paradoxical sleep
(b) Results of deafferentations
The possibility that a reflex mechanism with a peripheral point of departure is hlvolved in the production of a P.S. reflex should not be excluded a priori, since P.S. can be obtained by proprioceptive stimulation in pontile animals. Furthermore, the part played by the carotid sinus has often been referred to since the work of Koch (1932). Finally, changes in blood pressure (Candia et al., 1962) and in cardiac and respiratory rates allow for the hypothesis that P.S. is induced during slow sleep by means of nervous afferents with a vascular point of departure. In order therefore to rule out the possibility of exclusive triggering of P.S. by some extracerebral nervous mechanism we systematically eliminated the majority of nervous afferents (Fig. 17).
The following operations were carried out alone or successively in 10 normal cats: total section in two stages of the sino-aortic nerve, bilateral stellectomy verified by the appearance of a bilateral Claude Bernard-Horner's syndrome, bilateral vagotomy in the neck, and intradural section of the posterior nerve roots from C1 to C6.
The results, which were in the main negative, need not be reported in detail. None of the operations carried out alone produced appreciable EEG or behavioural changes during slow sleep or P.S. The average proportion of P.S. was not significantly different postoperatively. In 2 animals, in addition, total section of the sino-aortic nerve was successively followed by bilateral stellectomy and bilateral vagotomy, carried out in several stages. No great differences in slow sleep or P.S. were observed here either. Finally, it is noteworthy that suppression of nuchal EMG activity during P.S. was absolute in the 2 animals that had undergone section of the posterior cervical roots.
Six dogs exhibited completely normal slow sleep and P.S. on the criteria of EEG, behaviour, and the duration, after the fol]owing operations: total section of the spinal cord at C6, total medullary destruction from D5 to S2 together with bilateral section of the brachial plexus, abdominal sympathectomy, bilateral splanchnicectomy together with medullo-adrenal curettage, and total thyroidectomy.
These results are represented diagrammatically in Fig. 17. As will be seen, exclusive involvement in the inception of P.S. can be ruled out for the spinal cord below C6, the cervical sympathetic chain, the sinus and aortic nerves, the vagus, and, lastly, the posterior roots from C1 to C6. Participation of medullo-adrenal and thyroid hormones can also be excluded.
Mechanisms of paradoxical sleep
(c) Role of the hypothalamus and pituitary
The role of the hypothalamus has often been mentioned in the induction of sleep (Von Economo, 1929; Nauta, 1946) and endocrine influences arising in the hypothalamus or pituitary cannot be exc]uded a priori. The hormonal dependence of P.S. has, furthermore, been reported in rabbits (Faure, 1962; Kawakami and Sawyer, 1962. We therefore undertook a study of the possible role of the hypothalamus and pituitary. After total intercollicular section of the brain stem of 9 cats all the structures rostral to the section were removed, including the hypothalamus and pituitary, by curettage of the sella turcica (Fig. 18). The first group of 6 animals received no substitution therapy. Urinalysis showed polyuria with low specific gravity, a considerable reduction in sodium and an increase in potassium content. These animals all died after 6 or 7 days, presenting polypnea, tachycardia, hyperexcitability (starting at the slightest sound). Nevertheless, typical and regular periods of P.S. appeared during the first 5 days (Fig. 20). Their incidence declined regularly after the first 3 days (Fig. 19), but periods of 5 min were still registered 120 h after the operation. Brief periods of atony lasting 2-3 sec, accompanied by a few pontine spikes still appeared periodically on the 6th or 7th day. The last sign of P.S. to disappear was thus the monophasic pontine spikes, which were then no longer accompanied by complete disappearance of muscular tonus (Fig. 20). Substitution therapy (2 units total postpituitary extract daily, 1-2 units ACTH every second day) was started in 4 animals, either from the beginning or from the 4th day on. This therapy prolonged survival up to 1 month. P.S. reappeared regularly and periodically as in animals with an intact hypothalamic island. Thus, the appearance of P.S. more than 120 h after ablation of the hypothalamus and pituitary makes it possible to rule out the hypothalamic neurohormones and pituitary hormones as necessary criteria for the periodic incidence of P.S., for it can be presumed that 24 h after these ablations the hormones are no longer present in the blood.
Mechanisms of paradoxical sleep
(d) Deprivation of P.S. in the pontile animal
Since P.S. is the only form of sleep in pontile animals we turned to a study of the effects of deprivation. Our technique was as follows: at the beginning of each phase of P.S. the fall in nuchal EMG activity, by the intermediary of an Oneirograph, switches on a stimulator which applies electric shocks to the animal's log. The strength of these shocks is regulated to arouse the animal, in which case the reappearance of EMG activity automatically cuts out the stimulation.
Results: Deprivation was maintained for a maximum of 8-9 h. It appeared that when P.S. is suppressed by a shock it tends to recur at ever shorter intervals. At first these intervals are similar to those between periods of P.S. in controls. but after some hours they become so short that 40 60 sec after being wakened by the shock the animal falls into a new phase of P.S. (Fig. 21). Thus the total number of incipient phases of P.S. was as much as 155 in sessions that lasted for 9 h, and P.S. can hardly be suppressed for longer periods than this in view of the very great number of shocks required. When the stimuli were stopped, P.S. returned immediately. Its average duration was then slightly more than that of controls (7 as against 6 min), and its periodicity remained at a higher level for 2-3 h and then returned to normal.
A result such as this exemplifies the necessary nature of P.S. in pontile animals, it suggests an active process situated in the brain stem, the efficacy of which increases as its effect is suppressed.
Mechanisms of paradoxical sleep
(e) Effects of temperature on P.S. in the pontile animal
The poikilothermic pontile animal is subject to changes in the environmental temperature, so that the effects of hypothermia on P.S. can be readily studied. The fall in rectal temperature occurring when the animal is placed in a cold environment causes P.S. to disappear almost entirely. If, however, cooling is stopped and the rectal temperature remains stable, P.S. reappears periodically. Its duration is then in inverse proportion to the rectal temperature (Fig. 22). An increase in the duration of P.S. during hypothermia goes hand in hand with an increase in the length of the intervals between each episode of P.S., the proportion of the latter always remaining in the region of 10%. We have observed periods of P.S. lasting 25 min at a rectal temperature of 29°. At this temperature the animals exhibit generalized clonism of the head and limbs, even when they are returned to a warm environment. There is no real shivering, however. During phases of P.S. clonism disappears almost completely. If the animal is rewarmed so that the rectal temperature increases P.S. occurs more frequently and in animals whose temperature rises from 30° to 37° in 4 h the amount of sleep can increase to up to 35% of the total time.
In contrast, we have never observed P.S. at temperatures above 40.5�. (This was the maximum temperature attempted, as hyperthermia has serious effects in pontile animals.)
Mechanisms of paradoxical sleep
(f) Action of gamma-butyrolactone (G.B.L.)
The induction of P.S. by G.B.L. in intact or decorticated animals has already been reported (M. Jouvet, et al., 1961). lt is impossible, on the other hand, to produce P.S. by means of G.B.L. in animals bearing lesions of the pontine reticular formation, and this led us to suggest that the drug might act at the level of the pons. This hypothesis was confirmed by the observation of the action of G.B.L. in pontile animals possessing the hypothalamic island. Intraperitoneal injection of 50 mg/kg G.B.L. produced, after a latent period of 5-10 min the appearance of P.S. of which the behavioural and EEG aspects were identical with spontaneous P.S. The frequency of periods of P.S. is enhanced for 60-80 min, the proportion of P. S. rising to 30% during the hour following the injection. Higher doses of G.B.L. (100-200 mg/kg) produce a state of anesthesia, with effacement of EMG activity and no monophasic spikes or eye movements.
Mechanisms of paradoxical sleep
(g) Osmolarity of the blood and paradoxical sleep
Two observations led us to investigate the relationship between the periodicity of P.S. and changes in the osmolarity of the blood. Firstly it appears that copious forcing of fluids (120-160 ml) causes P.S. to disappear for several hours; we therefore administered 60 ml twice daily. Secondly, we had observed that certain states of dehydration (in the transitory diabetes insipidus following transection of the brain stem leaving a hypothalamic island) are accompanied by a marked, transitory increase in P.S., up to 20-30% of the total time. Fig. 23 shows the results of our investigations on the relationship between the osmolarity of the blood and P.S. in pontile cats with hypothalamic island.
(i) Hypo-osmolarity of the b]ood was obtained by drip infusion of a quantity of tepid water equalling 10% of the body weight, via a stomach tube over 30-60 min, together with injection of 1-2 units of an antidiuretic hormone. This treatment completely suppressed P. S. for 6- 10 h. P. S. then returned in short (1-2 min), infrequent episodes. It was found that dilatation of the stomach by the same quantity of air did not affect the rhythm or duration of P.S.
(ii) Hyper-osmolarity was obtained either by comp]ete withdrawal of liquid for 24 h and forcing an equivalent quantity of dehydrated food, or by intravenous injection of 20 ml of 20% hypertonic saline. In the latter instance the duration and frequency of P.S. increased a]most immediate]y and remained high for 5-6 h. When liquids were withheld the increase in P.S. was maintained for 24 h, but as the state of the animal deteriorated with continued dehydration (polypnea) the phases of P.S. diminished in duration and frequency and final]y disappeared. Resumption of normal hydration after 24 h was accompanied by an immediate return to norma] of the P.S. rhythm. A curious phenomenon was observed in all cases during the first phases of P.S. following resumption of a liquid diet: the appearance of a very regular rhythmic activity of 3-4 c/s in the pontine reticular formation (Fig. 24), first in bursts of several seconds associated with the pontine spikes, and then continuously until the reappearance of EMG activity. Such a pattern has never been observed in the waking state.
Discussion
(a) Duality of the states of sleep
The first section will be limited to positive evidence of the duality of the states of sleep and thus of the specificity and autonomy of paradoxical sleep in relation to slow sleep.
(i) Structural duality of the two states of sleep
It is difficult to explain the totality of the results we have just described on the basis of a 'unitary' theory of behavioural sleep, according to which similar mechanisms and identical structures are responsible for the two states of sleep. To explain the desynchronization of P.S. following synchronization of slow sleep in accordance with this theory, it has been necessary to presume the existence of a single progressively ascending inhibitory process, which attacks first the ascending activating reticular system and secondarily the thalamic synchronizing structures (Hernandez Peon, 1963). In point of fact, the structural details we have described necessarily imply different nervous structures responsible for the two states of sleep. The results of experiments with section and coagulation all make it possible to localize the structures triggering off P.S. in the pons (M. Jouvet, 1961; Cadilhac and Passouant-Fontaine, 1962; Rossi et al. 1963). We have discussed elsewhere (M. Jouvet, 1962a) the different arguments in favour of a descending origin of slow sleep from the telencephalon. But the initiation of this slow activity has not been finally cleared up. Some results indicate that synchronizing structures which could be responsible for slow sleep are to be found in the medulla oblongata (Moruzzi, 1960; Magnes et al., 1961). It has been suggested ( Rossi et al., 1963) that these structures are situated immediately behind those responsible for P.S. (in the caudal part of the pontine reticular formation). This hypothesis does not however take into account the fact that coagulation directly behind the nucleus reticularis pontis caudalis at the level of the anterior part of the nucleus gigantocellularis does not suppress slow sleep. Also there is no intermediary phase of sleep between wakefulness and P.S. in pontile animals as shown by either EEG or behaviour. If the synchronizing structures were situated in the lower portion of the brain stem, at the level of the pons and the medulla oblongata, we would have to assume that they could produce no EEG or behavioural manifestations in pontile animals. Moreover, the ascending synchronizing influences would have to act on structures rostral to the pons to produce behavioural sleep of an intermediary type between wakefulness and P.S. without slow activity, as occurs in decorticate animals (M. Jouvet, 1962a), whilst the cortex would be necessary to produce slow subcortical activity in intact animals.
(ii) Duality of the mechanisms of the states of sleep (Fig. 25)
Like the structural findings, the EEG, phylogenetic and ontogenetic data speak for a duality of mechanisms. The results of cortical and subcortical recordings which show the fundamental difference between P.S. and slow sleep are in fact confirmed by other methods. Thus, the cortical D.C. potential (Dement, 1964; Wurtz, 1964) in cats shows a sudden negative shift at the beginning of P.S., following the positive shift of slow sleep. The variations in cerebral impedance ( Birzis and Tachibana, 1964) and cerebral blood flow ( Kanzow et al., 1962) are similar to those observed during wakefulness and contrary to those observed during slow sleep. Lastly, studies with electrodes have shown a sudden increase in cortical (Evarts, 1962) or reticular activity (Huttenlocher, 1961) compared with that of slow sleep.
Variations in both the autonomic and somatic spheres show that P.S. is indeed a state qualitatively different from slow sleep, for blood pressure drops suddenly at the beginning of the former, while there is little change in the course of the latter (Candia et al., 1962). Finally, the monosynaptic spinal reflexes change little during slow sleep, and disappear entirely in P.S. (Giaquinto et al., 1964). All the results reviewed here cannot be explained on the basis of a 'unitary' theory of sleep.
Such a theory also necessarily implies that slow sleep (considered as a phase of light sleep) precedes P.S. (deep sleep). If the hypothesis is to be accepted, there ought to be a parallelism between the phylogenic and ontogenic evolution of the two states of sleep. But this is not in fact the case. Whereas slow sleep appears to be a characteristic of all vertebrate species studied polygraphically up to the present time (from reptiles to mammals), P.S. does not appear to be related to slow sleep in their phylogenic evolution. What is more, the absence of P.S. in chelonians, and its very rudimentary form in birds, contrast with its constancy and relative importance in all mammals. The number of species so far studied is too small to permit definite conclusions to be drawn, but it does appear that P.S. occurs from birds upwards on the evolutionary scale. It may therefore be presumed that with its appearance a new function came into being which is not necessarily related to sleep, since it does not appear in reptiles.
Ontogenic aspects also make a differentiation between the two states of sleep possible. At birth, in fact, P.S. occurs relatively more often and is less dependent on slow sleep than in later life. Periods of sleep with rapid eye movements have also been observed in newborns (Roffwarg et al., 1963; Delange et al., 1961). These phases of sleep similar to P.S. can also follow immediately on wakefulness. Thus in the newborn mammal, slow sleep is not a prerequisite of P.S. Until now, sleep states have been investigated primarily in animals that are very immature at birth and it is therefore diffficult when analysing the greater amounts and autonomy of P.S. compared with slow sleep to know what to attribute to the immaturity of the nervous system and what to the conditions of early life - milk diet, etc. Nevertheless, one fact stands out: in the cat the average duration of P.S. is almost from the first identical with that in the adult animals. It is only by a reduction in frequency that the proportion of P.S. versus total sleep diminishes. It must therefore be assumed that the P.S. mechanism is already at birth what it will be later in the adult, whereas slow sleep requires further deve]opment. In other words, P.S. depends on an 'innate' mechanism, whereas the mechanism of slow sleep is acquired after birth.
Whilst in adulthood a behavioural distinction can be made polygraphically between the two states of sleep, the fact that slow sleep habitually precedes P.S. would permit the conclusion that the former is a prerequisite of the latter. But our technique of selective deprivation makes it very easy to separate them. Our results confirm those of Dement (1960) in humans and show that a need for P.S. appears as soon as an attempt is made to suppress it. This suggests that a particular function is connected with the appearance of P.S. It is interesting to note in this context that a cat deprived of P.S. for more than 72 h resembles a newborn kitten in its recuperative sleep. In both cases P.S. constitutes the greater part of behavioural sleep (up to 80% of the first hour of recuperative sleep), phasic phenomena and periodicity are very much increased and, especially, P.S. can follow immediately on wakefulness, without being preceded by a phase of slow sleep. P.S. following directly on a state of wakefulness has also been observed in adult man during narcoleptic attacks (Rechtschaffen et al., 1963). Thus under particular conditions - selective deprivation of P.S., narcolepsy�not yet fully understood, it is possible to distinguish between the mechanisms of P.S. and slow sleep in adults.
Discussion
(b) Mechanisms underlying the appearance of P.S.
We shall discuss here the question of reflex production of P.S., the effect of deprivation of P.S. in pontile animals and humoral factors.
(i) Reflex production of P.S.
Inhibition of nuchal tonus during P.S. is not dependent on the gamma system alone, since it persists after section of the posterior cervical roots of the intact animal. Since, furthermore, the cerebellum is intact in pontile animals, it would appear that production of P.S. as a reflex depends on the integrity of the pontine reticular formation (posterior part of the nucleus reticularis pontis oralis and anterior part of the N.R.P. caudalis). There is thus no direct inhibitory effect emanating from the bulbar reticular formation (situated at the level of the nucleus giganto-cellularis and nucleus reticularis ventralis (Magoun and Rhines, 1946; brodal, 1957), since P.S. can no longer be triggered by mechanisms distal to the mediopontine or retropontine areas. Participation of adrenalin secretion can also be ruled out, as P.S. can appear immediately after nociceptive stimulation and, furthermore, injection of adrenalin does not produce P.S. It also appears that the integrity of the mesencephalic tegmentum prevents reflex P.S. in mesencephalic animals, although P.S. still occurs spontaneously. Activation of the descending facilitatory system (Magoun, 1950) would thus preclude descending inhibition in mesencephalic animals. We did not analyse the nervous effects of the natural stimuli used in our experimentation, so that it is difficult to isolate the particular type of afferents responsible for triggering P.S. But we do know that the majority of afferents of group III project principally on to the mesencephalic tegmentum, while those of groups I and II project for the most part on to the pontine reticular formation (Pompeiano and Swett, 1963), so that the latter may be responsible for producing reflex P.S.
Thus, under certain conditions, P.S. can be produced by a reflex mechanism. A distinction must however be drawn between generalized atony, a veritable reflex cataplexy, without pontine spikes or 'spindles' and eye movements, and true P.S. The former state presents no refractory phase and can follow any proprioceptive, or nociceptive, stimulus when the pons has been transected at the nucleus reticularis pontis oralis level. Such cataplexy resembles the 'sudden postural collapse' described by Bard and Macht (1958). It has never been possible, on the other hand, to obtain an iterative form of P.S. Thus, the data invite the conclusion that there exist two different systems at the level of the pons. The first, which is not endowed with refractory period, is responsible for inhibition of muscular tonus (probably via the inhibitory bulbar reticular formation), and the second, which controls the first and presents a refractory period, is responsible for pontine spikes and 'spindles' (and probably the EEG phenomena in intact animals) and for the eye movements. This second system can be identified with the 'centre déclenchant' of P.S. (M. Jouvet, 1962a) for this is the level at which the refractory phase producing the periodicity of P.S. appears. Such a system may explain the different aspects of certain reflex cataplectic attacks in humans. The one type - cataplexy - narcolepsy - presents EEG, polygraphic and electromyographic tracings identical with those in s]eep with eye movements (personal observation), and the other cataplexy - is characterized by intact consciousness.
If, therefore, P.S. can be induced as a reflex under certain conditions, this does not appear to be the sole mechanism. Nevertheless, this hypothesis has been maintained by Lissak et al. (1962). These authors contend that inhibition of muscular tonus is the cause, and not the effect, of P.S. But several facts make it possible to preclude the exclusive activation of P.S. by peripheral muscular afferents.
(1) Cortical activation of P.S. often precedes the disappearance of muscular tonus in intact animals.
(2) In pontile animals (without hypothalamic island) long periods of total atony are not infrequently observed. No EMG activity in the neck or other muscles is then present. ]n spite of this complete atony, P.S. can regularly appear, recognizable by pontine EEG criteria, rapid eye movements and autonomic changes. Total muscular atony is thus not per se an inhibitory or facilitatory factor of P.S.
(3) Lastly, total section of the spinal cord at C6 with section of the brachial plexus, whereby a large number of afferents of muscular origin are suppressed, does not affect the appearance of P.S.
Thus, the result of nervous deafferentation experiments is negative. Furthermore, the periodic and regular appearance of P.S. 4 5 days after ablation of the hypothalamus and pituitary rules out a direct and indispensable participation of ACTH and the post-pituitary hormones and the different hypothalamic neurohormones. The decrease in P.S. seen from about the fourth day and before death must be ascribed to delayed ionic and metabolic disturbances resulting from the prolonged absence of pituitary (especially antidiuretic) hormones.
The periodic appearance of P.S. thus cannot be explained by peripheral nervous or hypothalamo-pituitary factors, so that investigation can be restricted to pontine phenomena (neuronal or glial), although peripheral humoral factors cannot be excluded a priori.
(ii) It is difficult with the methods of study employed to specify the mechanisms of P.S. precisely. But several indirect findings all point to this state of sleep being endowed with a certain 'autorhythmicity' and situated in the pons.
The presence of P.S. under conditions of hypothermia at 29° indicates the resistance to cold of its underlying mechanisms. The findings are similar to those in anesthesia: P.S. in fact continues to appear periodically in decorticated animals after doses of 20-30 mg/kg pentobarbitone (M. Jouvet, 1962a). Furthermore, the greater duration of P.S. (and of the intervals between the phases) during hypothermia is reminiscent of enzymatic processes and suggests a metabolic phenomenon.
The action of y-butyrolactone (G.B.L.) is difficult to interpret, in spite of the fact that its structure is simple. Bessman and Skolnik (1964) see G.B.L. as a normal constituent of the brain, an increase in the intracerebral concentration coinciding with anesthesia induced by G.B.L. or gamma-hydroxybutyrate (G.H.B.). Giarman and Roth (1964), on the other hand, found no G.B.L. in the blood or brain and attribute the anesthetic role to y-hydroxybutyrate. G.B.L. does not appear to be a precursor of cerebral GABA, for the level of the latter does not increase after injection of G.B.L ( Giarman and Schmidt, 1963). It has also been shown that injection of G.B.L. can raise the level of acetylcholine in the brain (Giarman and Schmidt, 1963), and specifically in the area of the corpora quadrigemina. Whilst the mechanism behind this increase in cerebral acetylcholine is not known the ata suggest a cholinergic-type link within the P.S. process. Certain peripheral manifestations of P.S. in intact animals (myosis, bradycardia, increased and irregular respiration, arterial hypotension (Candia et al., 1962) inhibitory role of atropine (M. Jouvet, 1962a), support this view. But while such observations suggest the intervention of cholinergic neurons during P.S., they cannot explain the periodicity.
(iii) Deprivation of P.S.
The progressive diminution of the intervals between each incipient phase of P.S. in pontile cats caused by the increasingly frequent repetition of the disturbing shock, suggests a biochemical process whereby the accumulation during wakefulness of an unknown factor among the metabolites of neuronal activity provokes the increasingly rapid induction of a recuperative process. The inescapable nature of this reappearance of P.S. meant that with our technique prolonged deprivation was impossible. It nevertheless appears that a minimal increase in the recuperative process is adequate to restore the earlier conditions and normal rhythm of P.S.
In the intact animal, on the other hand, in which prolonged periods of deprivation are possible, P.S. increases considerably but does not exceed a certain ceiling (60%) during the first 6 h of recuperative sleep. This plateau thus appears to represent the upper limit of the cyclic metabolic processes of P.S. In this connection, it is interesting to note that recuperation does not occur in one long phase of P.S., but the periodicity remains, as though it were impossible for P.S. to last for more than 20-25 min at a stretch. Thus, the P.S. deficit which is accumulated over a long period of deprivation can only be partially and slowly compensated by a phenomenon which remains periodical and is of limited duration. This suggests the existence of a self-regulating cyclic metabolic process which requires several days before it can 'neutralize' the unknown factor accumulated during deprivation.
(iv) It is a difficult task to interpret the relationships between osmolarity of the blood and P.S., for the amounts of ions normally present in the cerebrospinal fluid and the brain are not known. The post-pituitary hormone could be cited as an indirect factor in the increase of P.S. during withdrawal of liquids and following injection of hypertonic saline solution. But injection of ADH does not promote P.S., which can occur in transitory diabetes insipidus (i.e. in the absence of post-pituitary hormones). There must therefore be another mechanism at work, which cou]d be none other than a direct influence of the osmolarity of the blood. In the case of pontile animals with an intact hypothalamic island, the relationship between the blood and the brain must always be considered in particular at the level of the barrier between the capillaries and the neuroglial cells, to which an important and active role is accredited in what is termed the blood-brain barrier (Edström, 1964). According to recent findings (De Robertis and Gerschenfeld, 1961), the neuroglia appears to be the tissue that plays the greatest part in the mechanisms of ion and water exchange between the internal milieu and the neurons. It constitutes a water ion pool (Gerschenfeld et al., 1959) between the blood and the neurons, playing a part in the transportation of metabolites and the storage or elimination of K and Na at the neuronal level. An active barrier, which has been likened to the renal glomeruli (Tschirgt, 1958; Edström, 1964), would thus keep the whole neuronal complex relatively independent of variations in the iOI1 content of the extracellular fluid. Furthermore, the osmolarity of the brain is closely related to that of the blood and quickly adjusts itself to variations in the latter (Stern and Coxon, 1964). There are thus grounds for supposing that variations in the osmolarity of the blood above all affect the glial cells. lt is therefore possible that an electro]yte concentration exceeding that of these cells facilitates the enzymatic mechanisms responsible for P.S., whereas a lower concentration would inhibit them. Lastly, the appearance of regular rhythmic activity during rehydration (after fluid deprivation) only during P.S. would appear to signify a close relationship between the electrical activity of the pons in P.S. and the osmolarity of the blood. One might therefore presume that at this moment, and only at this moment, the glial structures participate in the recurrence of a certain type of cerebral homeostasis. This hypothetical periodic glial mechanism, with its purpose either of maintaining cerebral homeostasis (cerebrostasis) or as an active process for the elimination of certain metabolites, would protect the brain so that P.S. would appear as a periodic phenomenon interrupting waking activity in the pontile animal and slow sleep in the intact animal, to restore equilibrium at the neuronal level. It must be admitted that if such a mechanism exists its location in the pons would be ideal. For, at that level, the nervous and glial cells could receive biochemical information on the activity of the neurons from the whole nervous system, since the afferents from the rostral and caudal regions of the nervous system converge at that point (Brodal, 1957), whilst efferent ascending and descending pathways lead out from the pons to the cortex and spinal cow (Scheibel and Scheibel, 1957).
(v) The nature of P.S.
Finally, we must gather the facts together and attempt to draw up some hypotheses on the nature of P.S.
It would appear to be established that P.S. is dependent on a periodic mechanism of an unknown nature situated in the pons. This mechanism triggers an ascending tonic neuronal activity similar to (or perhaps identical with) that of the waking state and a phasic activity which is specific to it. However intense the cortical, reticular or pyramidal neuronal activity during P.S., it is prevented from expressing itself (except for the rapid eye movements) in appropriate tonic motor phenomena by the inhibitory reticular formation set in action by the pons. The findings we have discussed also show P.S. to be a state differing qualitatively from slow sleep. The relation between the two is nonetheless a close one, however, for slow sleop is normally a precondition for P.S. Finally, numerous observations in humans show that dreaming is the subjective equivalent of P.S. (Dement and Kleitman, 1957; M. Jouvet and D. Jouvet, 1964).
Passing from the facts to hypothetical considerations, a number of points can be made. Recollection of recent or past events during P.S. suggests that certain memory processes occur during this phase. The relatively high incidence of P.S. in the neonatal period, when the 'plastic' processes of learning are at their most active, is also indicative of a probable relationship between P.S. and the process of memory. A further sign is the presence of a particular rhythmic 6-activity in the limbic system. The relationship of such activity during wakefulness to storage of information in the CNS has been discussed by Adey (1964). Finally the parallelism between variations in P.S. and the osmolarity of the blood suggests the intervention of the neuroglia, perhaps as regulator of certain processes of protein synthesis essential to storage in the neurons; this is in keeping with the findings of Hydén and Pigon (1960).
Is it possible that the biological clock in the pons which activates P.S. may be the mechanism responsible for the complex biological processes by which we retain (or lose) the memory of past events during dreaming ? It must certainly be admitted that if molecular changes are to occur at the level of the sensory and motor neurons it would be logical for them to occur during sleep and at a moment when a safety mechanism prevents peripheral motor expression of the discharges due to protein synthesis within the neurons, for otherwise the dreamer would run the risk of behaviourally reacting to dangerous hallucinations.
Paradoxical sleep, the physiological substratum of dreaming, would thus appear as the expression of a periodic function of storing information at the molecular level. Why this mechanism expresses a need, as objectified by deprivation experiments, remains to be explained. Are we to imagine a threshold in the functional information storage processes occurring in the waking state which initiates slow sleep? A large quantity of information (external stimuli, prolonged low-frequency stimuli) does in fact quickly produce slow sleep in the intact animal (supraliminal inhibition of the Pavlovian type, hypnogenic stimulation). It is possible that, in the pons, where the majority of neurons of the central nervous system converge, certain cells are subject to a biochemical mechanism which represents a certain threshold of functional storage of information. Once this threshold has been passed, a pontine mechanism would initiate the storage process at the molecular level. It is not unreasonable to presume that such processes are cyclic and autoregulatory and that they cannot exceed a certain limit of activity of 60%, as is the case during recuperation following deprivation of paradoxical sleep.
Summary and Conclusions
In the first part of this study, the arguments supporting the theory of the duality of sleep (slow sleep-paradoxical sleep) are set forth.
(1) Both tonic and phasic EEG or peripheral index of P.S. are totally different from EEG and behavioural slow sleep. The pontine origin of rapid eye movements and of the phasic ponto-geniculo-occipItal. activity occurring during P.S. is emphasized.
(2) Phylogenetic study shows that slow sleep may be observed in reptiles, birds and mammals. In contrast, P.S. is not found at all in the tortoise, and is of very short duration in birds (its ratio to the total sleep being only 0.2%). In mammals this ratio is about 6-30%).
(3) Ontogenetic studies in the kitten show that P.S. may appear immediately after wakefulness and constitutes 90% of total sleep during the first days. During maturation, the relative percentage of P.S. decreases to 30% while the percentage of slow sleep increases to 70% of the total sleep.
(4) The results of selective deprivation of P.S. in the adult animal are summarized. They show that after deprivation for more than 72 h, a maximum of 60% of P.S. is reached during recuperative sleep. This percentage is not exceeded even after 17 days of deprivation. On the other hand, during recuperation P.S. may be observed immediately after waking. After prolonged deprivation several days are required before the animal recovers the control level of P.S.
(5) Coagulation of the pons may suppress P.S. electively without producing any change in slow sleep.
(6) In chronic pontile animals, with hypothalamic islands, the rhombencephalic phase of sleep, showing all the pontine electrical and behavioural criteria of P.S. in the intact animal, can be completely identified with the latter. Its mean duration (6 min) is analogous to that of the intact animal while its duration per 24 h is somewhat less (10%). No behavioural or EEG index of slow sleep was observed in pontile animals.
All these results cannot be explained by a unitary theory of sleep. On the contrary, they allow us to differentiate P.S. from slow sleep in its structural bases and mechanisms.
The second part of the paper outlines some mechanisms of triggering P.S. in pontile animals.
It is possible to trigger reflex P.S. by proprioceptive or nociceptive stimulation in pontile animals, provided that the section is made caudally to the mesencephalic tegmentum. Various nervous deafferentations (afferents going through the spinal cord below C6, the vagus, buffer nerves) do not prevent P.S. from occurring in intact cats, and it does not therefore seem possible that P.S. is only triggered by reflex nervous pathways.
Ablation of the pituitary and hypothalamus does not suppress P.S. during the first 5 days of survival. A hormonal hypothalamo-pituitary mechanism is thus ruled out.
Deprivation of P.S. by electric shocks in pontile animals involves the reappearance, with increasing rapidity, of P.S., so that after a few hours P.S. can recommence several times per minute. This fact speaks for an active mechanism at the level of the pons.
The action of hypothermia on P.S. is considered and the resistance of this phenomenon to hypothermia shown.
The facilitatory effect of gamma-butyrolactone is emphasized. It was also found that P.S. is suppressed by hypo-osmolarity and facilitated by hyper-osmolarity of the blood.
All these results speak in favour of a self-regulating metabolic process, located in the pons, and the possibility of a neuroglial mechanism is considered.
Discussion
TISSOT : One form of sleep or several? The discussion brings to mind the classic quarrel between the neoticians and the associationists. The problem must be put differently. Although there are different structures, mechanisms and functions, often even opposed to one another, normal sleep has the characteristic quality of integrating them into a harmonious activity.
JOUVET: I agree with you, but if I adopt this way of trying to show the duality of the states of sleep it is because there is often a tendency to consider sleep as a single state with only quantitative changes - light or deep sleep. I think these terms are very misleading because they might give the impression that paradoxical sleep is only due to an intensification of the hypnic process whereas the occurrence of something else is in fact involved.
SOULAIRAC : The phylogenetic study of sleep ought to provide information about the physiological significance of the two types of sleep. The fact that only the higher vertebrates (mammals and to a very slight extent birds) manifest paradoxical sleep prompts the question whether it is not connected with the presence of the neocortical system.
One might suggest that just as development of the neocortex involves the appearance of a second vigilance system, superimposed on the fundamental mesencephalic system, paradoxical sleep would in the same way represent the manifestation of a second system of sleep regulation. This would then represent not a true duality but the coordination on the phylogenic plane of two chronologically different mechanisms controlling the unitary biological function of sleep in the higher vertebrates.
JOUVET : It would nevertheless seem that the cortex is not necessary for the occurrence of paradoxical sleep since the latter persists in decorticated or pontile animals. We also found to our surprise that, contrary to the classic theory, ontogenetic development does not simply repeat on a smaller scale the pattern of phylogeny since paradoxical sleep is particularly developed at birth, in both the kitten and the newborn human infant, whereas it is practically absent in birds. In this sense paradoxical sleep does not appear to be archisleop as we first thought.
HERNANDEZ-PEON : I share Prof. Hess' opinion supporting a unitary concept of sleep. Sleop and wakefulness are physiological states subserved by the dominance of antagonistic neural systems which must have a complex but integrated organization. I also agree with Prof. Jouvet that the region of the nucleus reticularis pontis caudalis plays an important role in the muscular hypotonia of the desynchronized phase of sleep. However, there is no need to postulate two different independent neural systems in order to explain the two groups of epiphenomena of sleep. Their habitual chronological relationship can be easily explained by different degrees of activation within a single sleep system which produces different degrees and extents of inhibition within the vigilance system.
JOUVET : I am afraid I do not agree with you at all. The fact that first slow then paradoxical sleep can be produced by injecting acetylcholine in situ is no proof that the same mechanism is responsible for both. Paradoxical sleep occurs spontaneously during slow sleep and it would be very naïve to imagine that it had been specifically produced when it occurs more than a minute after an injection. (The same applies, by the way, to slow sleep.) On the contrary we have demonstrated that slow sleep was not necessary for the occurrence of the paradoxical phase (ontogenesis, deprivation, pontile animals).
TISSOT : Dementia in advanced age has been shown to produce a longer duration of paradoxical sleep (as in the child) than that of the adult. This is another argument in favour of Jouvet's concept of archisleep.
JOUVET : I do not think we should generalize yet on the data concerning insanity. We have made tracings of psychotic subjects with Korsakoff's syndrome and have been surprised to see a significant decrease of paradoxical sleep in them.
MORUZZI : Prof. Jouvet has just given us new and very important facts on the phylogenic and ontogenic aspects of desynchronized or paradoxical sleep. I should like to ask his opinion of the relationship of classic or synchronized sleep to these bursts of desynchronized sleep which are of fairly short duration and characterized by essentially phasic phenomena.
What is the reason for the sudden disappearance of a phase of desynchronized sleep? An answer to this question might help us understand the functional significance of the phenomenon. Are we dealing with something new, which only interrupts synchronized sleep but which is not related to it and which disappears immediately and spontaneously, as any convulsive or subconvulsive manifestation? Or does synchronized sleep inevitably lead to paradoxical sleep and suppress it again as soon as it can deal with the situation?
It would be interesting to try to prolong the phases of paradoxical sleep in cases where it is very short, such as in the pigeon. A study of the postural and ocular effects in the thalamic pigeon would provide an answer to this question.
JOUVET : I think we are not yet in a position to solve this important problem. There seems to be no connection between the duration of slow sleep and that of the paradoxical sleep that follows. On the other hand, in about 70% of cases paradoxical sleep terminates in waking, however brief, and not in a return to slow sleep. We must regard paradoxical sleep as a self-regulated process of metabolic or enzymatic nature. Even after very long deprivation (22 days), such as we have recently carried out, the mean duration of recuperative paradoxical sleep falls to 6 min after 24 h. On the other hand, the intervals between each phase are shortened for some 12 days.
We have the impression that if paradoxical sleep represents either the synthesis or the elimination (or both) of some substance, this synthesis can only take place according to a process comprising a mechanism for its own autoregulation. The only way to increase the duration of paradoxical sleep (in the pontile animal) is hypothermia followed by rewarming.
It would of course be interesting to study thalamic pigeons, but I am afraid the phases of paradoxical sleep in birds are very short indeed.
KUGLER : Jouvet has demonstrated that in the case of the cat, the EEG patterns of certain phases of sleep accompanied by muscular atony are difficult to distinguish from the waking EEG. Hence he has coined the term 'paradoxical sleep' for these phases. In man, sleep accompanied by muscular atony and eye movements produces lowvoltage, fast EEG patterns instead of the high-amplitude slow activity of classic sleep. But they are not identical with the waking rhythm in humans. The frequencies are generally slower than those of the normal a-rhythm and present superimposed flat theta-waves. The patterns can more easily be compared with those of stage B of classic sleep. When alpha-like activity occurs it has a different topographical distribution and a different reactivity from the occipItal. alpha-rhythm. A characteristic of these phases, moreover, is the moment of occurrence: they occur only after classic sleep phases and not immediately after waking periods.
MINKOWSKI : I am wondering whether paradoxical sleep, accompanied by eve movements beneath closed eye-lids, as described so admirably by Prof. Jouvet, does not represent a recurrence - mutatis mutandis, of course - of the fetal form of sleep in the adult. Eye movements are very probably produced in the fetus long before the palpebral slits are formed, either as movements accompanying the first fetal head movements or as movements of proprioceptive origin in the strict sense of the word, i.e. produced by proprioceptive stimulation in the eyebal]s. From the anatomical aspect there is good reason to stress that the posterior longitudinal bundle in the protuberantial and mesencephalic tegmentum, one part of which unites and coordinates the oculomotor nuclei, is one of the formations that undergo myelinization earliest in the fetus - in the 4th month of intrauterine development - and that part of the fibres of the reticular formation and the nuclei and intracerebral parts of the oculomotor nerves undergo the same process shortly after. These formations thus constitute an anatomical fetal substrate for early eye movements which occur well before their postnatal reactions to visual stimulation.
As a result of my research into the successive development of nervous function in the early stages of the fetus, the newborn, the infant, the child and the adult, I formed the idea that each different phase of development does not completely disappear when the next one begins but continues to exist, at least potentially, at deep functional levels, and to interact according to the circumstances with the superimposed more highly differentiated adult components. It would be extremely interesting if this sleep could be demonstrated as manifesting the coexistence of elements of different genetic levels, the paradoxical phase of sleep appearing as a relic of fetal sleep in the general and complex sleep of the animal and adult human.
JOUVET : I fully agree with this interpretation. I think that experiments on apes will soon have reached the stage where we can study fetal sleep. The only facts we have so far refer to birds. Klein in my laboratory studied the chick embryo (from the 17th to the 20th day of incubation) and noted periods of rapid eye movements accompanied by bradycardia, but the cerebral electrical activity did not vary and no muscular activity could be recorded. These embryonic states possibly represent something connected with paradoxical sleep.
MONNIER : What differences in autonomic function have you observed during the two phases of sleep? How do the pupils react during paradoxical sleep?
JOUVET : The pupil is generally in a state of extreme miosis during paradoxical sleep (more contracted than during slow sleep). But short periods of mydriasis often accompany the sudden eye movements and even persist after cervical sympathectomy.
KONZETT : What is the effect of minimum doses of hypnotic substances on paradoxical sleep?
JOUVET : This question is very interesting. We have particularly studied the action of 'Nembutal'. This drug, even in anesthetic doses (30-35 mg/kg body weight), does not appear to suppress the occurrence of the paradoxical phase during anesthesia. Paradoxical sleep is certainly much more difficult to recognize than in the normal animal, but if the monophasic peaks in the pons or the lateral geniculate are taken as a criterion they will be seen to persist for regular periods of about 6 min during anesthesia and that their incidence is increased after selective deprivation. The process responsible for paradoxical sleep seems to be particularly active since it is not uppressed by anesthetic doses of 'Nembutal'.
PLETSCHER : What is the physiological significance of paradoxical sleep ?
JOUVET : I wish I could answer your question! This is certainly one of the most fascinating problems of the physiology of sleep. It would seem that paradoxical sleep may represent the expression of a 'dreaming function' and that it might possibly also have some connection with the phenomena of memory formation. A very speculative hypothesis would be that paradoxical sleep might represent the molecular synthesis of the proteins responsible for memory storage. This would explain the particular importance of paradoxical sleep at an early age when the learning processes are very much to the fore. We have equally been struck, as I mentioned just now, by the decrease of paradoxical sleep in subjects suffering from Korsakoff's syndrome (unpublished observations). However, I must confess that such observations are much too limited to enable us to establish a connection between paradoxical sleep and memory.
ARNOLD : Phylogenetically, sleep is a very old behavioural pattern. For this reason we are justified in discussing briefly the phenomenon of human sleep from the phylogenetic point of view. An important fact is that the development of man has been characterized by two absolutely opposed sleeping patterns. The earlier stage of phylogenetic development is represented by vegetarian tree-dwellers who also slept in trees. For this purpose the extremities must be kept in a cramped position (clinging function), which is only possible through a more or less upright position of the body produced by the corresponding tonus systems of the brain stem and spinal cord. In this sleeping position the distance between each member of the group is larger and the mutual protection smaller; defence or flight in the event of attack is only possible when signals (e.g. noise or vibration), perceived in spite of sleep, lead to immediate defence reactions.
At a later period, during the transition to prairie dwellers, quite different sleeping positions emerged with the development of a hunting community. While some kept watch the others huddled close together and lay in positions where the exposed surface of the body was reduced to a minimum as protection against loss of warmth and getting wet (fetal position). This demanded complete relaxation of the whole muscular apparatus, a horizontal position and a corresponding functional modification in postural reflexes.
These two phylogenically pre-formed and contrasting sleeping positions are still the extremes of a range of behavioural possibilities. Between these two extremes is a middle zone, which we might call the intermediary position. This perhaps includes paradoxical sleep with its numerous levels of activity expressed as frequent change of positions, muscular twitching and even sudden starts, attentive listening, getting up and sleep-walking.
It appears necessary not only to consider the complex pattern of sleep in man from the aspect of organization and fonction of neurophysiological substrates and their elements but also to bear in mind that the overall and detailed function of these elements is subordinate to the very old pattern of sleep and has wndergone the same phylogenic modifications.
Figure 1 : Polygraphic aspects of the two states of sleep

(A) Wakefulness: Fast cortical and subcortical activity.
(B) Slow sleep: Cortical and subcortical spindles and slow waves. Persistence of nuchal EMG activity (EMG). No eye movements (EYES).
(C) P.S.: Fast cortical activity similar to (A). Regular alpha-activity in the ventral hippocampus (HIPP). Phasic activity in the pontine reticular formation (PRF). Complete disappearance of nuchal EMG activity and rapid eye movements. Changes in respiratory activity (RESP) and the plethysmographic index (PLETH).
SMC = sensorimotor cortex; ESC = ectosylvian cortex. MRF = midbrain reticular formation.
Scale: 1 sec; 50 microV
From Jouvet,1962a.
Figure 2 : Phasic phenomena during P.S.

Normal cat: 3 days after enucleation of both eyes. Monophasic peaks grouped in pseudo-spindles in the pontine reticular formation (PRF), the oculomotor nucleus (N. III), the lateral geniculate nucleus (Lat. gen.) and the visual cortex (Vis. Cx.). Note phasic twitching of the whiskers, the temporal muscles, and the minor muscles of the face, and absence of nuchal EMG activity.
Scale: 1 sec; 50 microV.
Figure 3 : Electro-oculographic aspect of eye movements during P.S.

(a) Normal cat; (b) Pontile cat; (c) After coagulation of the superior colliculi; (d) After occipItal. decortication; (e) After total decortication. The oculographic tracings were made one month after the lesions.
Scale: 1 sec; 50 microV.
From Jeannerod et al., 1965.
Figure 4 : Persistence of phasic electrical activity in the visual system after enucleation of the eyeballs

(A) Phasic spikes during P.S. in the pontine reticular formation (PRF), lateral geniculate (Lat. gen.) and visual cortex (Vis. cx), with concurrent eye movements (EOG) and EMG activity from the m. rectus oculi internus (EMG m. rect. oc. int.).
(B) Four days after enucleation of the eyeballs. Persistence of phasic phenomena during P.S.
From Michel et al., 1964.
Figure 5 : Slow sleep and paradoxical sleep in the hen

(1) Wakefulness: fast activity in the hyperstriatum (EEG).
(2) Slow sleep: slow, high-voltage activity in the hyperstriatum. No true spindles.
(3) Paradoxical sleep (lasting 8 sec): considerable reduction, but not complete disappearance, of nuchal EMG activity; fast, low-voltage cerebral activity; note the burst of eye movements (EM). Bradycardia is also present.
Scale: 1 sec; 50 microV.
From Klein et al., 1964.
Figure 6 : Periodicity of P.S. in the chick

Chick aged 36 h. Diagrammatic representation of recording lasting 83 min. The black rectangles represent periods of P.S., which never lasted for more than 15 sec and were accompanied by a considerable reduction in nuchal EMG activity and by eye movements (+). The proportion of P.S. is 0.3%. Sleeping: 82.4%; awake 17.3%. Time scale: 1 min. Below: Changes in pulse rate (bradycardia) during the same period. The crosses represent P.S.
Time scale (abscissa): 1 min. Ordinate scale: pulse rate per min.
From Klein, 1963.
Figure 7 : Development of periodicity of P.S. in the kitten

Each circle represents 1 h; the black sectors show the average duration of phases of P.S., and the white sectors the intervals between. Key: (A) First week; (B) Second week; (C) First month; (D) Second month; (E) Third month; (F) Adult.
From Valatx et al., 1964.
Figure 8 : Diagram of the postnatal development of the states of sleep in the kitten

(Based on three-hour recordings daily.) Stippled: Wakefulness. Vertical hatching: Slow sleep with nuchal EMG activity. Crossed area: Slow sleep without tonic muscular activity. Diagonal hatching: P.S. The limit between the vertical white columns and the continuous marking represents the age at which cortical activity takes on the appearance seen in adulthood.
Abscissa: Relative proportion (%) of each state. Ordinate: Age in days.
From Valatx et al., 1964.
Figure 9 : Recuperation of paradoxical sleep after selective instrumental deprivation
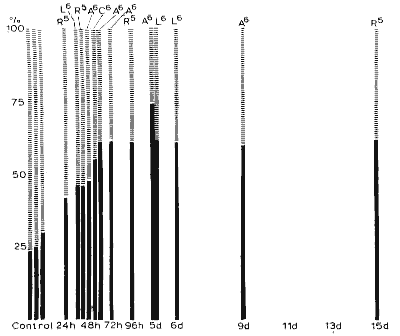
Ordinate: Black columns: paradoxical sleep in % of total sleep. Hatched columns: slow sleep. Based on the first 6 h of recuperative sleep after deprivation. Abscissa: Duration of P.S. deprivation inh (up to 96) and days. Data obtained in 4 cats. Note the remarkable constancy of the relative proportion of P.S. (60%) following deprivation of over 72 h.
Figure 10 : Differential effects of lesions of the lower brain stem on the two states of sleep

Lesions projected onto a sagittal section of the brain stem. Diagonal hatching: coagulation of the median three-quarters of the nucleus reticularis pontis oralis and interpeduncular nucleus. Persistence of P.S. (15%). lncrease in slow activity to 90% during first week. Return to normal proportion of slow sleep (60%) after third week (mean of 3 cats). Black area: lesion causing complete suppression of P.S. for 15-97 days. Mean proportion of slow sleep in 4 animals: 40%. Dotted area: Lesion in the caudal part of the nucleus reticularis pontis caudalis and the anterior part of the nucleus giganto-cellularis (involving the median two-thirds of these nuclei). Suppression of P.S. for 3 days followed by reappearance with normal rhythm (18%). Mean proportion of slow sleep in 2 cats: 40%. The proportion of slow sleep was determined in all animals in an average of 6 h of recording daily.
Figure 11 : Medio-sagittal section of the brain of a pontile animal with hypothalamic island

Anteriorly the trace of the brain stem section can be seen where the strip of acrylic resin was placed, and posteriorly the trace of an electrode in the caudal part of the nucleus reticularis pontis caudalis. Animal sacrified on 66th day. Luxol staining.
Figure 12 : Apparatus for the study of sleep in chronic pontile animals

The animal is placed in an incubator and its temperature maintained at a constant level (see details in the text). To obtain automatic deprivation of paradoxical sleep a system of relays (G) activates a stimulator (J) which delivers electric shocks to the cat's paw. The stimulation is interrupted on the resumption of nuchal EMG activity.
From M. Jouvet et al., 1963.
Figure 13 : Periodicity of P.S. in the pontile animal

Continuous recording with an Oneirograph for 5 days. Each 1ine represents alternately recording from 10 a.m. to 10 p.m. and 10 p.m. to 10 a.m. P.S. is represented by the vertical rectangles. The arrows show the times of forced feeding. Recording from 30th-35th day.
Time scale: 1 h.
Figure 14 : Pontine animal with hypothalamic island
 |
|
 |
|
 |
|
 |
EEG recorded 89 days after operation. The four tracings represent uninterrupted consecutive activity. Above: Arousal followed by paradoxical sleep with typical monophasic spikes at the pontine level (PRF), eye movements (EM) and total depression of muscular activity in the neck (EMG). The fourth tracing represents.the end of paradoxical sleep. Each line represents 2 min of recording.
Calibration 6 sec, 50 microV.
Figure 15 : Section of brain stem and reflex production of P.S.

Sagittal section of brain stem and location of the diflferent sections allowing or suppressing production of P.S. by proprioceptive or nociceptive stimulation. Black area: common lesion suppressing P.S. in intact animals. See text for details.
Figure 16 : P.S. and reflex cataplexy in the pontile animal

(1) Nociceptive stimulation of the ear (horizontal line) produces P.S. characterized by extinction of nuchal EMG activity and the appearance of a characteristic rhythmic activity in the nucleus reticularis pontis caudalis (PRF). Duration of reflex P.S.: 6 min;
(2) 3 min after cessation of P.S. the same nociceptive stimulation produces extinction of nuchal EMG activity for 3 min, but there is no change in activity in the pons.
Scale: 3 sec; 50 microV.
Figure 17 : Diagram showing deafferentation procedures that do not affect P.S.

(a) Ablation of the cerebellum, midbrain, hypothalamus, pituitary, diencephalon, and telencephalon. (b) Section of the posterior cervical roots from Cl to C6. (c) Section of the spinal cord at C6. (d) Section of the two vagus nerves and of the sino-aortic nerves. (e) Ablation of the two stellate ganglia. (f) Splanchnicectomy and medullo-adrenal curettage.
From D. Jouvet, 1962.
Figure 18 : Sagittal section of the brain stem of a pontile animal without hypothalamic island

Site of the electrode in the posterior part of the nucleus reticularis pontis caudalis. Loyez staining.
Figure 19 : Proportion of P.S. in pontile animals with and without hypothalamic island

Ordinate: Percentage of P.S. per 24 h (calculated from continuous Oneirograph recordings). Abscissa: Time in days. Continuous line: Pontile animals without hypothalamic island or substitutive therapy. Regular fall in P.S. and complete disappearance by 7th day (mean of 6 animals). Crosses: Pontile animal without hypothalamic island. The arrow shows the beginning of substitutive therapy (postpituitary extract and ACTH). Proportion of P.S. returns to normal by 15th day (mean of 3 animals). Dotted line: Pontile animal with hypothalamic island. A relative decrease in P.S. occurs about the 7th day. The average proportion of P.S. is 10% (mean of 15 animals).
Figure 20 : Development of pontine and EMG activity during P.S. in the pontine animal without hypothalamus or pituitary

- (a) 2nd day: monophasic pontine peaks during P.S.
- (b) 3rd day.
- (c) 6th day: muscular activity has not entirely disappeared and occurs phasically only during pontine rhythmic activity.
Scale: 1 sec; 50 microV.
Figure 21 : Automatic deprivation of P.S. in the pontile animal

Slow-speed recording. Disappearance of EMG activity in the neck on incipient P.S. produces an electric shock (artefact) which wakes the animal for some 10 sec until P.S. inevitably sets in once again (5th h of deprivation). PRF: Activity of the pontine reticular formation. Note the appearance of eye movements (EM) as EMG activity falls.
Scale: 3 sec; 50 microV.
Figure 22 : Effect of temp rature on the rhythmicity of P.S.

Ordinate: Time in min. Abscissa: Rectal temperature. Dashed line: Average duration of P.S. at different temperatures. Continuous line: Average interval between episodes of P.S. Mean of 3 animals.
Figure 23 : Relationship between P.S. and osmolarity of the blood

Pontile animal with hypothalamic island. Ordinate: Proportion of P.S. during recording period (calculated per period of 100 min). Abscissa: Time in minutes and hours. The normal proportion of P.S. (control determined over 10 days) is 8% (+ 2%). Continuous line: After intravenous injection (arrow) of 20 ml of 20% hypertonic saline, high levels of P.S. for 5 h. Broken line: After forcing of a quantity of water 10% of the body weight, following injection of 2 units ADH. complete disappearance of P.S. for 5 h followed by slow return to normal. Blood Na and K levels were determined at A, B, C and D (in mequiv/l): before and after injection of saline: Na: 142-152-168-157; K: 6.4 5.2-6.3 6.2. a b c-d : after forcing of water: Na: 157-146-138-136-138; K: 6.2-6.6-6.2-5.6 7.2. In this animal injection of ADH had no effect on the proportion of P.S.
Figure 24 : Relationship between pontine electrical activity during P.S. and the osmolarity of the blood

Chronic pontile animal with hypothalamic island. (1) P.S. during dehydration. Monophasic spikes grouped in pseudospindles, accompanied by lateral eye movements (EM). The pontine electrode is situated in the nucleus reticularis pontis caudalis. (2) P.S. after rehydration (60 ml liquid). Very regular theta rhythm of 4.5 c/s associated with spikes and eye movements. This activity was observed only during the first two phases of P.S. after rehydration. It was never observed in the waking state. Identical pontine activity has been observed in 4 animals under the same conditions.
Figure 25 : Duality of the two states of sleep

Ordinates, for each diagram: percentage of P.S. (in black) with respect to behavioural sleep.(A)Phylogenetic aspect: hatched: newborn; in black: adult animal. (1) Tortoise: no P.S.; (2) Hen (0.2%); (3) Lamb (17%); sheep (6.6%); (4) Young rat (55%); rat (15%); (5) Kitten (80%); cat (30%); (6) Infant (50%); man (20%). From Klein, 1963. (B) Ontogenetic aspect (in kittens). Abscissa: Age in days. From Valatx, 1963. (C) After selective deprivation of P.S. during recuperative sleep. Abscissa: Duration of deprivation in days. (D) After destruction of the pontine reticular formation (posterior part of the nucleus reticularis pontis oralis and anterior part of N.R.P. caudalis. No P.S. (E) Chronic pontile animal. No slow sleep.