| Le sommeil paradoxal : Est-il le gardien de l'individuation psychologique ? |
| Michel Jouvet Revue Canadienne de Psychologie, 1991, 45(2), 148-168 |
| TABLE DES MATIERES |
| IMPRESSION |
| Version imprimable (Tout l'article dans une seule page) |
Modalités synchroniques
Les modalités théoriques d'une programmation génétique du système nerveux central
Neurogenèse : Il est évident qu'au cours de l'ontogenèse, la neurogenèse en contribuant à l'organisation génétiquement programmée du système nerveux central est la gardienne de l'individuation. Ainsi, une neurogenèse qui persisterait au cours de la vie adulte pourrait continuer à assurer cette fonction.
Chez les poissons, amphibiens et reptiles (poïkilothermes ou ectothermes), il existe une croissance continue du cerveau pendant toute la vie. Celle-ci est assurée par une neurogenèse continuelle au cours de la vie post-natale (voir revue in Holder & Clarke, 1988). Ainsi, chez les poissons, il peut exister un remplacement continuel de certaines cellules olfactives dont les axones vont rejoindre les neurones centraux. Cette neurogenèse contribue à garder l'information de l'imprinting pendant toute une vie en permettant le homing du saumon (Hasler & Scholz, 1983). Chez les amphibiens, une neurogenèse continuelle peut exister également au niveau du système rétinotectal. Chez les reptiles (lézards), une neurogenèse post-natale a également été démontrée au niveau du cortex cérébral (Lopez-Garcia, Molowny, Garcia-Verdugo, & Ferrer, 1988). Il semble exister enfin une corrélation entre l'existence d'une neurogenèse continuelle et la faculté de régénérescence du système nerveux puisque la moelle épinière caudale peut entièrement être reconstituée à partir de cellules périépendymaires chez les amniotes (Holder & Clarke, 1988).
L'apparition de l'homéothermie, en permettant une liberté plus grande par rapport aux conditions thermiques de l'environnement, s'est accompagnée d'une diminution considérable des capacités de neurogenèse post-natale. Les oiseaux semblent représenter une étape transitionnelle: le cerveau des oiseaux possède en effet des similarités importantes avec celui des poissons, amphibiens et reptiles (voir revue dans Konishi, Emlen, Ricklefs, & Wingfield, 1989). De nouveaux neurones peuvent apparaître au niveau du cerveau antérieur des oiseaux adultes. Ils effectuent une migration depuis leur lieu de naissance juxtraventriculaire jusqu'à leur destination en glissant sur la piste des cellules gliales radiées (comme chez les ectotherrnes). Dans certains cas, ces nouveaux neurones peuvent être incorporés dans des circuits fonctionnels (par exemple le système responsable du chant chez le canari ou le pinson (Zebra Finch) (Nordeen & Nordeen, 1990; Paton & Nottebohm, 1984). Il semble que ces processus de neurogenèse adulte apparaissent de façon saisonnière en relation avec les stéroïdes sexuels. Ainsi, il peut exister chez les oiseaux des possibilités de neurogenèse adulte qui pourraient contribuer à entretenir une programmation génétique.
Par contre, ces phénomènes n'existent pas chez les mammiferes chez qui toute neurogenèse disparaît rapidement au cours du mois qui suit la naissance. Au lieu d'une neurogenèse, c'est au contraire une régression qui apparaît avec la mort des périkaryas et la rétraction des axones au cours de la vie adulte. Cet aspect régressif s'accompagne également d'une diminution considérable des capacités de régénérescence des neurones centraux (voir revue dans Oppenheim, 1985). Seuls les neurones olfactifs sont continuellement remplacés au cours de toute la vie (voir références dans Farbman, 1990). Les pistes des cellules gliales radiées qui assurent la migration des neurones au cours de l'ontogenèse disparaissent également chez les mammifères.
En conclusion, une conservation de l'individuation génétique par neurogenèse permanente chez l'adulte semble probable chez les ectothermes (poissons, amphibiens et reptiles). Elle est également possible, mais saisonnière, chez certains oiseaux adultes. Elle est totalement impossible chez les mammifères après la période post natale. Il faut donc supposer qu'un nouveau mode de programmation génétique soit apparu avec l'homéothermie.
La programmation itérative: La programmation des circuits responsables de l'idiosyncrasie héréditaire ne nécessite pas, en effet, l'apparition de nouveaux neurones par neurogenèse. Les périkaryas contiennent le DNA nécessaire à la synthèse des molécules qui sont incorporées dans les membranes comme récepteurs. On peut ainsi émettre l'hypothèse que certains types de neurones, qui apparaissent tardivement au cours de l'ontogenèse (comme les neurones Golgi de type II de Jacobson, 1970), puissent synthétiser continuellement des récepteurs. Ces récepteurs labiles ne deviendraient fonctionnels que s'ils étaient excités (stabilisés) par des excitations endogènes.
Il faut alors présumer qu'il existe un générateur endogène qui soit responsable de la validation de ces récepteurs selon le même mécanisme utilisé par les stimuli épigénétiques pour maintenir fonctionnelle la voie visuelle par exemple. Il n'est pas besoin d'admettre la nécessité d'un codage temporel d'influx à partir du générateur pour instruire l'entretien et la validation des systèmes responsables des réactions idiosyncrasiques héréditaires (Hypothèse Instructive, Jouvet, 1978). Selon une hypothèse sélective plus simple (Jouvet, 1980, 1986), l'information de la programmation héréditaire dépendrait alors seulement de la sélection des neurones qui sont, ou ne sont pas, excités par une programmation endogène dont le codage peut être stochastique. D'après cette hypothèse sélective, un nombre limité de gênes pourrait contribuer à la programmation de l'hérédité psychologique en induisant à la fois la synthèse des récepteurs et l'arrivée des terminales présynaptiques issues du générateur au niveau de différents groupes d'interneurones (voir Fig. 1).
Modèle théorique de programmation génétique itérative au cours du sommeil paradoxal
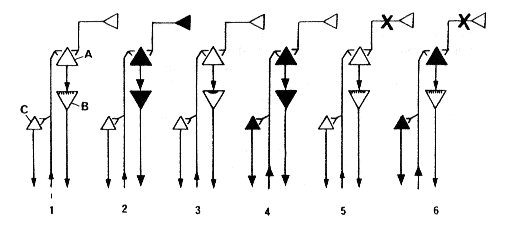
(1) Un neurone B (Golgi type II) vient de synthétiser un récepteur qui est incorporé dans la membrane sous forme labile (hachures verticales).
(2) Au cours du SP, le système PGO (en haut à droite) stimule l'interneurones A, ce qui va exciter et stabiliser le récepteur du neurone B et le rendre fonctionnel (comportement onirique).
(3) Ce récepteur demeure fonctionnel pendant l'éveil qui fait suite au SP.
(4) Aussi long temps que le récepteur est fonctionnel, les stimuli de l'environnement (flèche ascendante) peuvent activer à la fois des réponses non spécifiques d'éveil (système C) et la réponse idiosyncrasique génétiquement programmée à partir de B. Si le SP est supprimé
(5) et qu'il n'existe plus d'activité PGO, le récepteur reste labile, non fonctionnel, et le neurone B n'est plus activé.
(6) Dans ce cas, le stimulus épigénétique ne sera plus capable d'entraîner de réponses idiosyncrasiques, mais seulement une réponse non spécifique. Le chloramphénicol pourrait inhiber la synthèse du récepteur B et ainsi découpler l'activité PGO des réponses unitaires du neurone B et de son expression motrice dans le comportement onirique (voir détail dans le texte; d'après JOUVET, 1980).
Les mécanismes et les conditions théoriques d'une programmation génétique itérative : Le générateur endogène doit pouvoir à la fois agir au niveau des systèmes perceptifs et des systèmes moteurs corticaux et sous-corticaux. Dans le premier cas, il pourrait être responsable de l'induction de perceptions sans objet ou d'hallucinations. Dans le deuxième cas, il doit induire la maintenance ou la facilitation des programmes de comportement responsables des réactions idiosynchrasiques individuelles ou éventuellement supprimer ou altérer certains programmes moteurs acquis au cours de l'apprentissage. Il apparaît donc nécessaire que l'excitation programmée puisse agir jusqu'au niveau des motoneurones ponto-bulbaires ou médullaires.
Les motoneurones étant soumis à l'influence d'une programmation endogène, deux conséquences devraient en découler:
(1) Il devrait être possible, dans certaines conditions, de distinguer une composante génétique dans les patterns d'activité des motoneurones qui sont directement, ou indirectement, sous le contrôle du générateur au cours de la programmation. Cette composante génétique traduirait la mise en jeu de différents groupes d'interneurones en réponse à l'arrivée des excitations stochastiques délivrées par le générateur endogène.
(2) Il devrait également exister certains mécanismes capables de supprimer l'activité musculaire au cours de la programmation du système moteur ou des systèmes perceptifs sinon l'organisme serait soumis à de possibles hallucinations ou à des comportements moteurs stéréotypés non contrôlés qui risqueraient d'être dangereux s'ils survenaient au cours de l'éveil ou du sommeil. Afin que la programmation génétique puisse survenir sans trop de bruit de fond et que le rapport signal sur bruit soit élevé, il faudrait également qu'il puisse exister des mécanismes de contrôle qui inhiberaient l'arrivée des influx sensoriels extérieurs ou internes. Ainsi, les processus d'intégration seraient facilités au niveau des systèmes programmés (comme cela se produit au cours de l'attention dans la vie éveillée).
Enfin, étant donné qu'au moment de la programmation génétique le cerveau ne peut recevoir et répondre aux stimuli extérieurs (et probablement à certains signaux internes), du fait de l'inhibition des afférences intéro- et extéroceptives et du blocage de l'activité musculaire, il devrait exister un mécanisme de protection qui permettrait à ce processus de ne survenir que lorsque l'organisme n'est plus soumis à des stimuli potentiellement dangereux, c'est-à-dire à un moment où les mécanismes d'éveil ne sont plus mis en jeu, c'est-à-dire au cours du sommeil.
Modalités diachroniques de la programmation itérative
La programmation génétique est évidemment prédominante au cours de l'ontogenèse, c'est-à-dire pendant la période d'organisation du système nerveux central par neurogenèse in utero ou in ovo. Ensuite, la durée de la programmation ne peut que diminuer car elle ne peut survenir qu'au cours du sommeil. On peut donc supposer que chez l'adulte il puisse exister un rapport entre la durée de la programmation et celle du sommeil.
On doit supposer que la programmation du système nerveux central nécessite et utilise une grande quantité d'énergie, au moins égale à celle qui est utilisée pendant l'éveil au cours de l'apprentissage. Des réserves énergétiques sont accumulées au cours du sommeil sous la forme d'une glycogénogenèse (Karnovsky, Reich, Anchors, & Burrows, 1983). Ces réserves pourraient procurer l'énergie nécessaire à la programmation. La meilleure solution serait alors de concevoir un fonctionnement périodique: chaque épisode de programmation serait suivi d'une période de renouvellement de réserve au cours du sommeil. Ainsi, la programmation itérative pourrait s'effectuer selon un mode périodique.
Afin d'être efficace, la programmation génétique périodique doit être en relation avec les stimuli épigénétiques qui ont excité le système nerveux central au cours de l'éveil précédent. C'est-à-dire qu'il devrait exister des mécanismes pouvant adapter la durée de la programmation à la quantité de stimuli, soit extérieurs, soit intérieurs (hormonaux), qui pourraient avoir induit des modifications épigénétiques au niveau des systèmes dépendant des synapses cibles de la programmation.
Cependant, la programmation génétique de la personnalité est aveugle sur le plan du destin de l'individu. Elle est la loi et n'a pas de raison de faciliter la trace de tous les événements qui ont excité le cerveau. Dans certains cas, sans doute, la programmation pourrait faciliter l'acquisition d'un apprentissage si celui-ci renforce certains traits idiosynchrasiques de la personnalité. Dans d'autres cas, il devrait n'avoir aucun effet ou même pourrait inhiber ou effacer certains circuits synaptiques épigénétiquement acquis si ceux-ci s'opposent à la typologie. Ce concept d'efface ment de certains circuits corticaux au cours du SP (Jouvet, 1980) a été particulière ment développé par Crick et Mitchinson (1983). Selon ces auteurs, la fonction du SP serait surtout de supprimer certaines modalités indésirables d'interaction au niveau du cortex. Ce processus, appelé reverse learning pourrait surtout servir à oublier: "we dream in order to forget". Ainsi, la suppression de la programmation génétique devrait soit supprimer, soit faciliter, ou n'avoir aucune action sur l'apprentissage. Par contre, on doit s'attendre à ce que cette suppression puisse altérer l'expression des comportements idiosynchrasiques et ainsi diminuer la variance phénotypique entre individus appartenant à des souches génétiques différentes au sein de la même espèce.