The Effects of Spinal or Mesencephalic Transections on Sleep
Related Erections and Ex-Copula Penile Reflexes in the Rat
Markus H. Schmidt, (MD, PhD)*, Kazuya Sakaï **, Jean-Louis Valatx**, and Michel Jouvet, (MD) **
Sleep . Vol 22 Number 4 pp 423-432 (1999)
** Université Claude Bernard, Departement de
Medecine Experimentale,8, Ave. Rockefeller, 693 73 Lyon, France
* Cleveland Clinic Foundation, Department of Neurology, 9500 Euclid Ave.,Cleveland,
Ohio 4419
Summary: The neural mechanisms of penile erections during paradoxical sleep (PS) remain unknown since it has yet to be the subject of neurophysiological investigation. Using a new experimental model for sleep-related erection research in freely behaving rats, neural transections were undertaken to definitively elucidate the effects of paraplegia on PS-related erections and to determine at which brain level the mechanisms underlying PS erectile activity are generated. Continuous polygraphic recordings, as well as ex-copula penile reflexes, were performed in male Sprague Dawley rats before and after spinal (n=4) or mesencephalic (n=6) transections. Spinal transections virtually eliminated PS-related erections. Following mesencephalic transections, PS remained qualitatively intact in all rats. PS erectile activity, however, was severely disrupted, as shown by a significant decrease in the total number of erections, the number of erections per hour, and the percentage of PS phases exhibiting an erectile event. Finally, spinal and mesencephalic transections had contrasting effects on ex-copula penile reflexes. Spinal transections significantly shortened the latency to reflex induction and increased the percentage of tests eliciting an erectile event, whereas mesencephalic transections significantly increased the latency to reflex induction without affecting the percentage of tests eliciting an erectile event. These data suggest that the brainstem is not sufficient for the generation of PS erectile activity even though it is sufficient for the generation of other classic PS phenomena. We conclude that neural structures rostral to the mesencephalon (i.e., the forebrain) are essential for the maintenance and integrity of PS related-erections. The reflex erection data suggest that spinal transection removes a tonic descending inhibition of erections, whereas such an inhibition not only remains intact, but appears enhanced following mesencephalic transection. We hypothesize that the forebrain plays a facilitatory role in erectile control, at least in part, through disinhibition of brainstern tonic anti-erectile mechanisms.
Key Words: Sleep-related erection; penile erection; erectile mechanisms; paradoxical sleep; REM sleep; nocturnal penile tumescence
INTRODUCTION
PENILE ERECTIONS ARE A CHARACTERISTIC PHENOMENON of paradoxical sleep (PS), or rapid eye movement (REM) sleep, in all normal human males from infancy to old age. (4,19) Since the discovery of PS-related erections in the mid 1960s, (4,19) sleep-related erection testing has served as a clinical tool to evaluate erectile dysfunction. The neural mechanisms of such erections, however, remain unknown since they have yet to be investigated. We recently demonstrated that rats also exhibit penile erections during PS, (38) thus establishing an animal model for sleep-related erection research. This new animal model provides the first opportunity to examine sleep-related erectile mechanisms and to answer fundamental questions regarding PS erectile control.
Penile erections involve the complex coordination of parasympathetic, sympathetic, and somatic motor components, incorporating neural elements spanning virtually all levels of the neuraxis. (6) These elements at the spinal level comprise what has been termed the spinal generator since erections may occur without descending input from higher brain structures. (6,31) Indeed, it has long been hypothesized that the brain exhibits a tonic descending inhibition since reflexive erections are facilitated by spinal transections (3) or spinal block. (29)
A lthough spinal mechanisms have been relatively well elucidated,6 the supraspinal control of penile erections is poorly understood with numerous structures from the medulla to the telencephalon hypothesized to play a role in erectile mechanisms. These structures include the medullary paragigantocellular nucleus (nPGi), (21,22) paraventricular nucleus, (25) medial preoptic areal, bed nucleus of the stria terminalis, (42) amygdala, (20) and olfactory bulb.(3) It is currently unknown whether these diverse structures may be involved in a single complex network controlling penile erections across multiple behavioral states, or whether these various structures are associated with multiple independent systems involved in the state-specific generation of penile erections.
Given the strong association of penile erections with PS, we sought to examine whether the neural mechanisms of these sleep-related erections are closely associated with the executive structures of this sleep state. It is now generally accepted that the executive mechanisms of PS are located within the brainstern since PS persists following mesencephalic transections (14) or even following the complete removal of all neural elements rostral to the pons, as in the "pontine cat" preparation. (15) The dorsal pontine tegmentum and rostral medulla contain the PS subsystems responsible for the various tonic and phasic phenomena characteristic of this sleep state, including cortical desynchronization, (13,41) rapid eye movements, (40) ponto-genicu lo-occipital (PGO) spikes, (32,40) and muscle atonia. (9,33) Whether the brainstern also is sufficient for the generation of PS-related erections, however, remains to be investigated.
Using a new animal model for sleep-related erection research, the neural mechanisms of PS-related erections were examined for the first time in the following experiments. First, midthoracic spinal cord transections were performed to definitively elucidate the effects of paraplegia on PS-related erections. Second, mesencephalic transections were undertaken as a first approach in localizing neural structures responsible for PS-related erections and to determine if the brainstern is sufficient for PS erectile genera tion. Finally, reflexive erections also were performed in both spinal and mesencephalic rats during wakefulness to examine the descending control of penile reflexes in the two groups. Preliminary results of these data were presented in abstract form. (34,35)
MATERIAL AND METHODS
Twelve male Sprague-Dawley rats (IFFA-CREDO, France) of 335-390g body weight were used for these experiments. The rats were maintained on a 12:12 hour light-dark cycle with food and water available ad libitum. The ambient temperature was maintained at a constant 25.0 ± 0.5°C.
Implantation
All rats were implanted for chronic penile erection monitoring and standard sleep recording under pentobarbital anesthesia (60 mg/kg). Penile erections were recorded using a technique previously described, (37,38) involving pressure monitoring within the bulb of the corpus spongiosurn penis (CSP) and simultaneous electromyography (EMG) of the ischiocavernosus (IC) and bulbospongiosus (BS) muscles. Briefly, a 3 French micro-tip pressure transducer (O.D.: 1.2 mm, Millar Instruments, Inc.) was implanted into the bulb of the CSP. The IC and BS muscles were then implanted with bipolar, Teflon-coated, hook electrodes (0.002 in. uncoated diameter, multistranded stainless steel wire, AM-Systems, Inc.) with the aid of a 20-ga hypodermic needle according to a method described by Holmes et al. (10)
Implantation for standard sleep recordings involved electrooculography (EOG), dorsal neck EMG and cortical electroencephalography (EEG). Electrical signals from the pressure catheter and the cable connected to the animal were passed through an electronic swivel system (Air Precision) to allow free movement of the animal following implantation.
Data Acquisition
Continuous polygraphic recordings were made from freely behaving rats with an ECEM polygraph at a paper speed of 5 mm/sec. Three 24-hour control recordings were made prior to transection, after which rats were recorded continuously for several weeks. Each rat, therefore, served as its own control.
Wakefulness, SWS, and PS were scored from 30-second epoclis using classical scoring criteria.26 In a "cerveau isolé" preparation cortical desynchronization cannot be used as a sign of PS since brainstern PS executive structures are physically separated from the forebrain. PS phenomena, therefore, must be examined caudal to the transection. PS in the "cerveau isolé" rat is defined as the simultaneous occurrence of muscle atonia and rapid eye movements, Wakefulness and SWS following mesencephalic transections were defined as non-PS episodes with cortical desynchronization or slow wave activity, respectively. Finally, the PS episode duration is defined as the duration of PS uninterrupted by either SWS or wakefulness. Any two PS episodes separated by less than four minutes in duration comprise a single PS phase. Polygraphic recordings were made for an average of 12.8 days following mesencephalic transections, with the longest recording of 24 days.
Erectile events were scored in relation to CSP pressure changes. That is, an erectile event was defined as a minimum increase of 30 mm Hg in CSP pressure with at least one pressure peak greater than 100 min Hg above the " flaccid baseline" level. The end of the erectile event was defined as the moment when the tumescent CSP pressure fell below the 30 min Hg threshold for more than 15 seconds. The following variables were then calculated for each 24-hour recording: 1) the total number of erections during each behavioral state; 2) the number of erections per hour of each behavioral state; and 3) the percentage of PS phases and episodes associated with at least one erectile event.
Penile erection monitoring using this technique simultaneously records micturition events.(36) These micturition events are associated with a distinct pattern of CSP pressure and perineal muscle activity that is entirely different from that seen during erectile events.36 During micturition, CSP pressures exhibit a rapid series of short duration (55 msec) pressure peaks at a frequency of 9-11 Hz throughout the micturition duration (3-5 sec). The number of micturition events during each behavioral state was systematically recorded.
Spinal Transection
Spinal transections were performed on four rats following control recordings. Under pentobarbital anesthesia (60 mg/kg), a 2-3 cm midline incision was made along the back over the midthoracic vertebrae. The muscles and overlying connective tissue were retracted and a laminectomy performed on the T8 vertebral segment. The spinal cord was then transected using a scalpel blade. Gelfoam was packed into the wound and the overlying muscles and skin sutured closed. The wound was cleaned with antiseptic solution, and a local antibiotic (Soframycine) was injected subcutaneously at the wound site.
Spinal rats were placed back into their cages immediately following the lesion. Post-transection care involved daily subcutaneous injections of gentamycin (2 mg). In addition, the bladder was voided twice daily by applying pressure to the lower abdomen, after which the perineum and lower abdomen were briefly cleaned. Bladder voiding by massage was continued until a return of spontaneous micturition was observed.
Mesencephalic Transection
In a separate experiment, mesencephalic brainstem transections were performed in eight rats. Following control recordings, rats were anesthetized with either pentobarbital (60 mg/kg) or Ketamine (80 mg/kg) and placed in a stereotaxic instrument. Mesencephalic transections were performed under stereotaxic guidance with a retractable wire knife. With the wire knife pushed out of the cannula, the wire tip, with respect to the cannula tip, was ventral (V) 2.5 mm and either L 5.0 min or L 6.0 min depending on which knife was used. The knife cannula was inserted in the brain at P 5.8 and at L 2.5 mm (for the 5.0 mm knife) or L 3.0 mm (for the 6.0 mm knife). The wire knife was pushed out of the cannula at V -2.0 to -3.4 mm with respect to the surface of the brain. The cannula was then lowered to V -8.5 to -9.0 mm at which point the wire knife was retracted and the cannula raised out of the brain.
Following mesencephalic transections, the animals were given daily subcutaneous injections of either isotonic saline or isotonic glucose with saline (5 to 7 ml) containing antibiotics (Gentamycin, 2 mg). The animals also were fed a fortified concentrated milk solution (5 to 7 ml) by esophageal intubation twice daily. These animals exhibited spontaneous micturition immediately following transection and, therefore, did not require bladder voiding by massage. Finally, rectal temperature was checked regularly after transection.
Reflexive Erections
Reflex erection tests were performed on six mesencephalic rats and three spinal rats before and after transections using a modified method of Sachs and Garinello.(10) The animals were placed on a board in a supine position with the head and anterior torso in a black Plexiglas cylinder. The abdomen was secured to the board with masking tape, allowing the head and anterior torso to move freely inside the cylinder. All rats were habituated to this procedure four times before implantation (2 x 5 minutes and 2 x 20 minutes). Reflexive erections were initiated by retracting the penile sheath and maintaining it in a retracted position by placing the glans penis through a hole in masking tape fastened to the abdomen. Once the penile sheath was retracted, the reflex test was continued for 20 minutes following the first penile response or stopped after 20 minutes if no erectile events were observed. Because of the delicate nature of the mesencephalic transections, only one or two reflex tests were performed on each rat before and after transection. Post-transection reflex tests for both spinal and mesencephalic rats were performed 7 to 13 days after the lesion
The following responses or variables were recorded during reflex erection tests: E1, weak glans engorgement involving the base of the glans; E2, moderate glans engorgement involving some dilatation of the distal glans; E3, intense flaring or cup of the distal glans; F1, dorsiflexion of the penile body; F2, dorsiflexion or flip of the penile body greater than 90 degrees with respect to rat's body; and, finally, the latency to the first erectile response following retraction of the penile sheath. An erectile event, or cluster, as it is defined in the literature (21,30) is a group of consecutive responses, glans erections, or flips, separated by less than 15 seconds. The intercluster interval (ICI) is the average time between clusters, calculated by dividing the time between the-end of the first cluster and the beginning of the last cluster by the total number of clusters in the reflex test.
Histology
Spinal rats were sacrificed at the end of the experiment by lethal injection of pentobarbital. The spinal cord was visually inspected and the level and completeness of the spinal transection verified by exposing the vertebral column.
Mesencephalic rats were sacrificed under deep pentobarbital anesthesia by transcardial perfusion of 250 ml Ringer's lactate solution (0.1% heparin) followed by a 500 ml ice-cold solution of 2% paraformaldehyde in a 0.1 M phosphate buffer. The brains were later sliced in the sagittal plane on a cryostat into 25 µm sections. Floating sections were mounted on gelatin-coated slides and stained with cresyl violet to analyze the extent of the transection. Lesions from sagittal sections were analyzed and reconstructed on a frontal plane to compare the extent of the transections among rats.
Data Analysis
Two 24-hour control recordings, as well as two post-transection recordings from the seventh to ninth day post-lesion, were used for data analyses. The average values for each variable recorded during sleep and wakefulness, spontaneous erections, and reflexive erections were calculated for each animal pre- and post-lesion. Paired t-tests and one way ANOVA's of the means of the means were employed for statistical analyses. For the ANOVA calculations, a post-hoc Tukey test was utilized on significant (p<0.05) F values. All statistical analyses were performed with Statgraphics (Manugistics) software on a PC computer.
RESULTS
Pre-Lesion Recordings
Control recordings prior to spinal or mesencephalic transections revealed a distinct relationship of penile erections with the animal's behavioral state. This state dependency of erectile activity was most apparent regarding the number of erections per hour of each behavioral state. As Figure 1 demonstrates, rats exhibited a mean of approximately four erections per hour of wakefulness, 10 erections per hour of PS, and a virtual absence of erections during SWS prior to spinal or mesencephalic lesions.
Spinal Transection
Spinal transections of all four rats were complete and located between T7 and T9 spinal cord segments. All animals were able to eat, drink, and move about in the cage with the use of the forelimbs following spinal transection at this level. Although the hindlimbs were completely paRalyzed, the withdrawal reflex in response to a paw pinch was well intact. Finally, the micturition reflex recovered in all rats within six to eight days post-transection. An analysis of these events revealed that the only significant change following the spontaneous return of micturition was the occasional micturition event occurring in PS and SWS, which was not observed in the control condition.
Table 1 : Analysis of paradoxical sleep, slow wave sleep, and wakefulness during a 24-hour period before and after spinal or mesencephalic transection
| Spinal Transection | Mesencephalic Transection | |||
| C | T | C | T | |
| Total PS (min) | 101 ± 12 | 81 ± 10 | 95 ± 2 | 54 ± 5 |
| Number of PS Phases | 45.5 ± 10 | 38.1 ± 5.4 | 45.8 ± 2.0 | 27.3 ± 3.1 |
| Avg. PS episode dur. (min) | 1.9 ± 0.1 | 1.7 ± 0.1 | 1.6 ± 0.1 | 1.8 ± 0.2 |
| Total SWS (min) | 639 ± 25 | 613 ± 27 | 673 ± 21 | 449 ± 30 * |
| Total Wake (min) | 683 ± 33 | 731 ± 37 | 664 ± 18 | 931 ± 35 * |
Note: Values are given as the means ± S.E.M. The data from the spinal and mesencephalic rats represent the mean values from two control recordings (C) and two post-transection recordings (T) obtained from the seventh to ninth day post-lesion. SWS and wakefulness following mesencephalic transection were defined as non-PS episodes with cortical desynchronization or slow wave activity, respectively. The PS episode duration is defined as the duration of PS uninterrupted by either SWS or wakefulness. Any two PS episodes separated by less than four minutes in duration comprise a single PS phase. * Indicates a significant difference with respect to the corresponding pre-lesioned control using paired t-tests; *p<0.001. C, control; T, post-transection.
Fig. 1: The number of erections per hour of wakefulness (W), paradoxical sleep (PS), and slow wave sleep (SWS) before (white) and after (shaded) spinal or mesencephalic transection.
* Indicates significant difference with respect to the prelesioned controls using paired t-test; *p<0.01, **p<0.001.
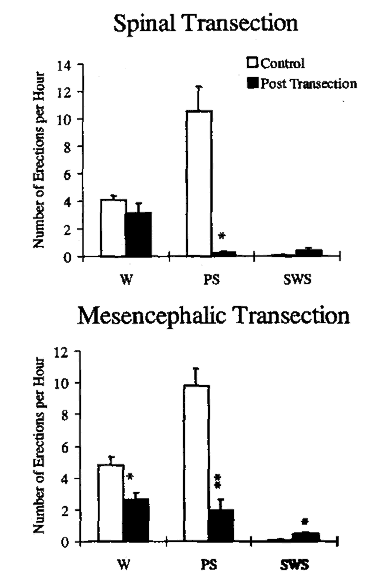
Table 2 - Analysis Or 24-hour erectile activity across behavioral states before and after spinal or mesencephalic transection.
| Spinal Transection | Mesencephalic Transection | |||
| C | T | C | T | |
| Paradoxical sleep | ||||
| Total Erections | 18.1 ± 4.0 | 0.4 ± 0.2 | 15.3 ± 1.7 | 1.7 ± 0.6 |
| Percent Phases with Erection | 36.5 ± 5.6 | 0.8 ± 0.5 | 31.2 ± 2.5 | 7.0 ± 3.0 |
| Slow Wave Sleep | ||||
| Total Erections | 0.6 ± 0.3 | 4.1 ± 2.0 | 0.9 ± 0.3 | 3.8 ± 0.8 |
| Wakefulness | ||||
| Total Erections | 46.5 ± 3.0 | 36.9 ± 7.0 | 52.8 ± 5.3 | 40.2 ± 7.4 |
Note: Values are given as the means ± S.E.M. The data from the spinal and mesencephalic rats represent the mean values from two control recordings (C) and two post-transection recordings (T) obtained from the seventh to ninth day post-lesion. * Indicates a significant difference with respect to the corresponding pre-lesioned control using paired t-tests; *p<0.05, **p<0.01, ***p<0.001. C, control; T, post-transection.
Although sleep-wake parameters remained unchanged post-lesion (Table 1), we observed a dramatic and significant reduction in PS erectile activity. The most striking effect was a significant reduction in the number of erections per hour of PS (Fig. 1). Similar reductions in PS erectile activity were found with respect to the total number of erections during PS, as well as the percentage of PS phases associated with an erection (Table 2).
Unlike the disruption in PS-related erections, the quantity of waking-state erections were only minimally decreased following spinal transection (Table 2 ). These erectile events during wakefulness, however, were related almost exclusively to movement of the animal such as during drinking, eating and, especially, during movement of the animal using the forelimbs, causing the perineurn to brush against the surface of the cage. When the animal was relatively motionless during quiet wakefulness or sleep, however, erectile events were rarely observed and appeared only sporadically during these behavioral states. During sleep, for example, post-hoc comparisons following a one-way ANOVA revealed that the number of erections per hour following spinal transection was not significantly different between PS and SWS.
Mesencephalic Transection
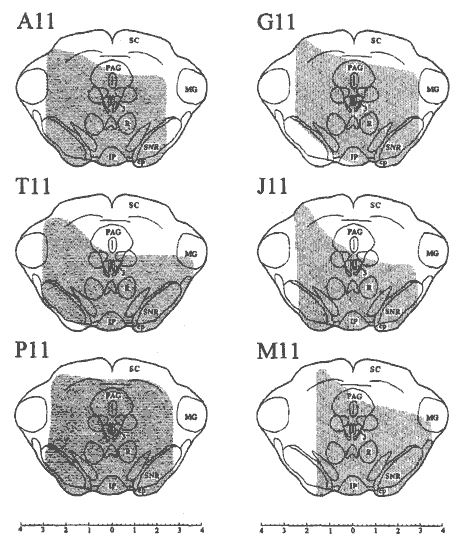
Two of the eight rats used for this study did not survive mesencephalic transection and are not included in the following data analysis. Mesencephalic transections of the remaining six rats were located at the same rostro-caudal level resulting in transection of the rostral superior colliculus and red nucleus (Fig. 2). Moreover, we found minimal rostro-caudal extension of the transection (Fig. 2). Exceptions included partial degeneration of both rostral and caudal portions of the transected red nucleus, as well as some hemorrhaging restricted to midline structures in several rats that often involved the oculomotor and Edinger-Westphal nuclei, but leaving the midline raphe nuclei intact in all cases. In all rats the medial geniculate nucleus, lateral extremes of both the substantia nigra and cerebral peduncles, and dorsal portions of the superior colliculi were not transected (Fig. 3). It should be noted that the completeness of the transections varied among rats. For example, the Periaqueductal Gray (PAG) remained largely intact in two cases (T 11 and J 11), and the lateral extent of the transection was not symmetrical in one rat (M 11) in that a large portion of the left central tegmental field, substantia nigra and cerebral peduncle was not transected (Fig. 3). These individual differences are highlighted when necessary when physiological effects differed.
Fig. 2 - Photomicrograph of two sagittal sections demonstrating the rostral mesencephalic transection of rat A11.
The approximate laterality (L) of the sections with respect to the midline are indicated in the figure according to the atlas of Paxinos and Watson. (28)
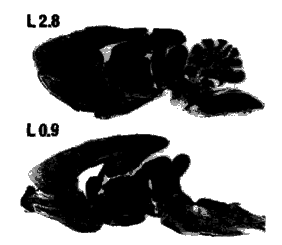
Fig. 3 - Frontal representations of the mesencephalic transections in six rats.
Lesions from sagittal sections were analyzed and reconstructed on a frontal plane, corresponding to P 5.8 on the Paxinos and Watson (28) atlas. SC, superior colliculus; PAG, periaqueductal gray; MG, medial geniculate nucleus; R, red nucleus; SNR, substantia nigra pars reticulata; IP, interpeduncular nucleus; cp, cerebral peduncle; 3, oculomotor nucleus. Scale at the bottom of the figure is in mm. The drawing of the coronal section was adapted from ref. (28)
Periodic monitoring of rectal temperature demonstrated that all rats were able to maintain a normal body temperature of approximately 37.0°C following transection. In addition, all rats maintained the micturition reflex immediately following the mesencephalic lesion. The only significant feature regarding an analysis of micturition was a 70 percent reduction in the total number of micturition events with more than a doubling in the duration of each event in these rats.
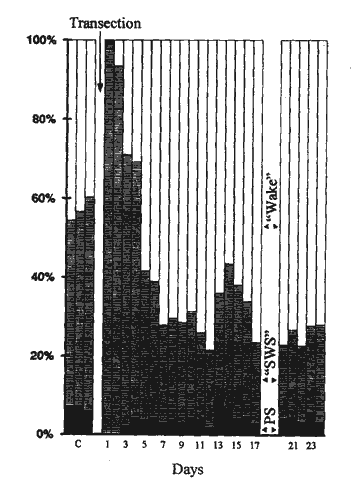
Following mesencephalic transection, PS phenomena were still present in all rats as seen by the rhythmic occurrence of muscle atonia and rapid eye movements (Fig. 4). These PS episodes were usually, though not always, associated with a cortical desynchronization. PS was generally absent for the first 24 hours following the transection, reappeared after two to three days, and the total daily quantity of PS stabilized within four to five days post-transection (Fig. 5). The total daily quantity of PS, however, was significantly reduced one week following transection (Table 1). This decrease in total PS was related to a reduction in the number of PS phases and not in the average PS episode duration (Table 1).
Cortical slow wave activity, or SWS, markedly increased for the first two to four days post-transection (Fig. 5) as previously demonstrated in the literature in cerveau isolé preparations. (8,27) Cortical desynchronization then reappeared and dominated the polygraphic recordings one week post-lesion. Indeed, following an initial recovery period, there were both a significant increase in wakefulness and a decrease in SWS one week post-transection with respect to control levels (Table 1).
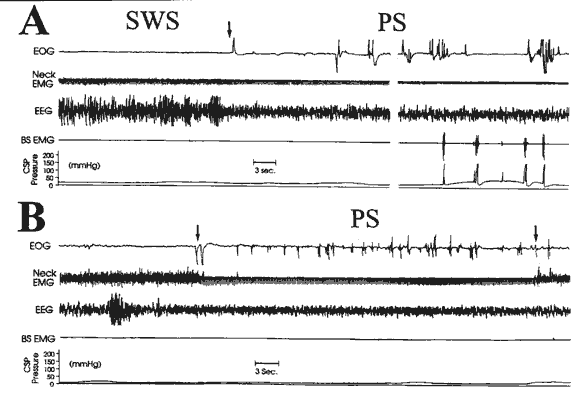
Fig. 4 - An example of a polygraphic recording demonstrating paradoxical sleep (PS) before (A) and after (B) mesencephalic transection.
A: An example of a transition from SWS to PS (see arrow) prior to mesencephalic transection and a typical erectile event during PS occurring approximately thirty seconds later in the same PS episode. PS-related erectile events involve a characteristic increase in baseline CSP pressure and, with BS muscle bursts, dramatic pressure peaks. B: PS episode following a mesencephalic transection in the same rat. Note the simultaneous occurrence of muscle atonia and rapid eye movements which define the PS episode. The arrows mark the beginning and end of the PS episode. Activity seen in the neck EMG during the PS episode is EKG artifact on an otherwise atonic baseline. The CSP pressure during PS post-transection typically remained stable at the flaccid baseline level with an absence of BS muscle bursts.
Although PS remained qualitatively intact, PS-related erections were disrupted following mesencephalic transections (Fig. 4). PS erectile activity was significantly reduced from an average of 9.8 erections per hour pre-lesion to less than two erections per hour of PS (Fig. 1), even after survival periods as long as 24 days post-transection. As shown in Table 2, this reduction in PS-related erectile activity post-transection also involved a highly significant decrease in the total number of PS-related erections and the percentage of PS phases exhibiting an erectile event.
The quantity of PS-related erections remaining following mesencephalic transection appeared to be related to the completeness of the transection. For example, in rat M I I the lateral extent of the transection was not bilaterally symmetrical (Fig. 3), leaving a large portion of the left rostral mesencephalon intact. This rat exhibited the most PS-related erectile activity following transection with an average of 5.0 erections per hour of PS more than one week post-lesion. In contrast, no PS-related erectile events were observed during the entire 12-day, post-transection, recording period of rat Al I in which the mesencephalic transection was relatively complete (Fig. 3).
We also found a significant decrease in the number of erections per hour of wakefulness (Fig. 1). This per hour reduction was related to the significant increase in total wakefulness following the transection since the total number of erections during this state did not significantly decrease (Table 2). On the other hand, SWS was associated with a small but significant increase in the number of erections per hour after transection.
Reflexive Erections
Control, or pre-transection, values from six mesencephalic rats and three spinal rats were pooled together for an analysis of penile reflexes. The ability to induce a reflexive erection was dependent on the level of the transection. For example, whereas 100% of all reflex tests were successful in eliciting erectile events following spinal transection, only 50% of all reflex tests following mesencephalic transection were successful in eliciting penile reflexes, identical to the control condition. Moreover, the latency to reflex induction was significantly decreased following spinal transection, whereas it was significantly increased post-mesencephalic lesion (Table 3). The number of erectile events, or clusters, tended to decrease following mesencephalic transection, but this reduction did not reach statistical significance at the p<0.05 level. Finally, neither spinal nor mesencephalic transection had a significant effect on the total number of glans erections (E1, E2, or E3), flips (F1, F2), or the inter cluster interval (Table 3).
Table 3 - Effects of Spinal or Mesencephalic transection on penile reflexes
| Latency (sec) | Total Glans Erections | Total Flips | Number of Clusters | lCl (sec) | |
| Control | 293 ± 58 | 24.0 ± 2.7 | 10.2 ± 2.3 | 11.1 ± 1.4 | 108 ± 16.7 |
| Spinal | 80 31 | 20.0±3.1 | 11.0±2.5 | 8.2±1.3 | 151 ±22.7 |
| Mesencephalic | 725 135 | 13.0 ± 6.1 | 8.5 ± 3.8 | 5.3 ± 2.2 | 127 ± 8.3 |
Note: Values represent the means ± S.E.M. Statistical analyses were performed using a one-way ANOVA El E2 and E3 events were combined as total glans erections, whereas F1 and F2 vents were combined as total flips since no sig- nificant differences in the number of these events were observed between groups. * indicates significant difference with respect to controls using a post-hoc Tukey test; *p<0.05, **p<0.01. ICI: intercluster interval.
Fig. 5 - The percentage of the total recording time spent in PS (black), SWS (shaded), and wakefulness (white) for three control recordings (C) and up to 24 days post-transection of rat P11.
Following mesencephalic transection, wakefulness, and SWS were defined as non-PS episodes associated with cortical desynchronization or slow wave activity, respectively.
DISCUSSION
This is the first report examining the neural mechanisms of penile erections across sleep-wake states. Using a new animal model for sleep-related erection research, we transected either the spinal cord or rostral mesencephalon in the rat to definitively elucidate the effects of paraplegia on sleep-related erections and to localize neural structures responsible for PS erectile generation. In addition, reflexive erections were performed in both spinal and mesencephalic rats to evaluate the descending control of penile reflexes in the two groups.
Spinal Erectile Mechanisms
Published data regarding the effects of paraplegia on human sleep-related erections are minimal, inconclusive, and contradictory. Although sleep-related erections are commonly thought to be disrupted by spinal cord injury as originally reported in two early abstracts, (17,18) Halstead and colleagues (7) later report in a full publication that human PS-related erections may persist following "complete" upper motor neuron spinal lesions. Any significant PS erectile activity remaining following complete upper spinal cord transection would suggest that the spinal generator controlling erections is activated or modulated by either a humoral component during PS or by simple peripheral reflex activation associated with phasic PS-related muscle twitching above the level of the transection. These latter data only confuse our understanding of sleep-related erection neurophysiology and require clarification or further exploration.
Our results demonstrate that complete midthoracic spinal transections virtually eliminated PS-related erections. Moreover, any remaining erectile activity observed during sleep was found to occur at random during SWS and PS, suggesting intermittent reflex activation of the spinal erection generator during sleep. Other sleep-wake parameters, in contrast, remained unchanged post-lesion. These data clearly demonstrate that an intact spinal cord above the midthoracic level is essential for the integrity of PS erectile activity. It remains to be determined, however, if PS erectile activity would be disrupted following spinal transections below the thoracolumbar level, leaving sympathetic innervation from the brain to the penis intact.
Although PS-related erections were adversely affected after spinal transection, spinal erectile mechanisms remained intact in all rats. Indeed, ex-copula reflexes following spinal transection demonstrated that erectile events were more easily elicited with shorter latencies relative to controls. These results confirm previous data demonstrating that reflexive erections are facilitated following spinal transection (31) or spinal block, (29) suggesting that spinal transection removes a tonic descending erectile inhibition.
In contrast to the decrease in PS-related erections, the total number of waking-state erections, as well as the number of erections per hour of wakefulness, did not significantly decrease following spinal transection. The variability in the waking-state erection data, however, markedly increased post-lesion. Moreover, these waking-state erections following spinal transection were associated almost exclusively with movement of the animal, especially when the perineurn would brush against the surface of the cage. We hypothesize that these waking-state erections were reflexively induced post-lesion. When the animal was relatively motionless during quiet wakefulness or sleep, erectile events were rarely observed after spinal transection.
Brainstem Mechanisms
Brainstem transections have served as an effective technique to localize neural structures responsible for PS generation. (14,43,44) Such data demonstrate that the pons and rostral medulla contain all of the neural elements necessary to generate the classic tonic and phasic phenomena characteristic of PS, (14) including cortical activation, rapid eye movements, muscle atonia, and PGO spike activity. A fundamental question is whether the brainstern also is sufficient to generate PS erectile activity.
Penile erections during PS were found to be dramatically and significantly decreased following mesencephalic transection even though PS was found to persist post-lesion. Moreover, the amount of PS erectile activity remaining post-lesion was related to the completeness of the transection, in that the most complete transections eliminated PS-related erections. These findings suggest that although brainstern PS mechanisms persist post-lesion, as seen by the rhythmic occurrence of rapid eye movements and muscle atonia, neural structures rostral to the transection (i.e., in the forebrain) are necessary for the production of PS-related erections.
Unlike its clear documentation in the cat, (14) PS has never been definitively demonstrated in the rat post-mesencephalic transection. (5,8,27) We observed a rhythmic occurrence of PS in all mesencephalic transected rats as seen by the simultaneous appearance of rapid eye movements and muscle atonia. PS was generally absent for the first 2-3 days post-transection, but stabilized in total daily quantiby day 4-5. Our greater success in observing PS in cerveau isolé rats may be secondary to the significantly longer survival periods in this study (average of 12.8 days vs 2-4 days in other reports). In addition, our transections were located at the mesencephalic-diencephalic junction which is more rostral than transections attempted in rats in previous reports. This more rostral location of the transections may potentially have resulted in less damage to midline pontine structures important for PS generation. Finally, although the transections were not complete as demonstrated histologically (Fig. 3) and polygraphically by the frequent onset of cortical desynchronization during PS, the transections were extensive enough to severely disrupt forebrain PS erectile mechanisms. It should also be noted that PS persisted even in rats with the most extensive transections when PS often was associated with cortical slowave activity.
Although PS remained qualitatively intact, we observed a significant reduction in the total daily quantity of PS one week post-transection. Since our previous results suggest that the probability of observing an erectile event during PS diminishes with decreasing PS episode duration, (38) it was essential to determine if the disruption in PS erectile activity was secondary to a simple reduction in the duration of PS episodes. Further analyses revealed, however, that the reduced total daily quantity of PS was characterized by a decrease in the total number of PS episodes without affecting the average, uninterrupted, PS episode duration.
Reflex erection tests following mesencephalic transection revealed that the latencies to induction were increased, and the percentage of tests eliciting an erectile event were similar to controls. These findings suggest that a descending inhibition not only remains intact but perhaps may be enhanced in a decerebrate preparation. Indeed, recent investigations have identified a major source of erectile inhibition within the brainstem in the rostral, juxtafacial, paragigantocellular nucleus (nPGi). (21,22) In the anesthetized rat, transection of the brainstern caudal to the nPGi removes a descending inhibition of penile reflexes comparable to spinal transection, whereas the inhibition remains intact following transections rostra] to this nucleus. (22) Moreover, bilateral electrolytic or cytotoxic lesions of the mPGi appear to facilitate reflexive erectile events. (21,22) Our reflex erection data demonstrating an intact descending inhibition following transection of the rostral mesencephalon supports the finding of a major inhibitory source penile erections within the brainstem.
Although our results suggest that the brainstem may be sufficient for the normal generation of PS-related erections, the data do not exclude a potential role of the brainstem in the direct descending control of PS erectile activity. For example, the nPGi appears to involve serotonin as a neurotransmitter in tonic descending erectile inhibition. (23) Given that brainstem serotonergic neurons cease firing during PS (PS-off neurons), one may speculate that a decrease in serotonergic inhibition from the brainstem could play a role in the production of PS erectile activity.
Following early transection experiments which localized the brainstem as the generator of PS, it has been assumed that the brainstem is the direct source of all PS-related phenomena. Recent experiments in the cat, however, demonstrate that mesencephalic transection eliminates phasic alterations of systemic arterial blood pressure which typically occur during PS in the intact animal (16). Together with our data, we hypothesize that certain PS phenomena, such as penile erections and PS-related cardiovascular changes, require forebrain structures. It remains to be determined, however, whether the forebrain mechanisms of both penile erections and cardiovascular control are related.
Forebrain Mechanisms
These experiments suggest that neural elements rostral to the mesencephalon are essential for the maintenance and integrity of PS-related erections, providing major directions for future research regarding the localization of neural structures responsible for PS erectile control. Unfortunately, it is presently difficult to speculate where this forebrain control may be derived. Numerous forebrain structures have been implicated in the control of either penile erections or sexual behavior and, therefore, are potential candidates in erectile mechanisms. These structures include the medial preoptic area (1,24,39) bed nucleus of the stria terminalis, (42) hippocampus, (2) amygdala, (20) and paraventricular nucleus. (25) The involvement of these structures in sleep-related erection physiology is unknown. We hypothesize that the preoptic, basal forebrain area may be a major candidate in sleep-related erectile control given its role in both sleep generation (12) and reproductive mechanisms. (1,6)
It remains to be determined how forebrain structures may influence brainstem or spinal erectile mechanisms during PS. We hypothesize that the facilitatory role of the forebrain in PS erectile control may include, at least in part, an inhibition of tonic brainstem inhibitory mechanisms. Indeed, our reflex erection data suggest that a descending erectile inhibition may be enhanced following mesencephalic transection. Such a disinhibitory role of the forebrain in erectile control has been suggested by others. (24)
Additional research is required to localize forebrain structures important for PS erectile generation and to elucidate the descending control of sleep-related erectile mechanisms. All available surgical or medical options for the treatment of erectile failure, including Sildenafil (Viagra), target the end organ level. Further understanding of higher central erectile mechanisms, Particularly forebrain PS erectile control, may revitalize and redirect sleep-related erection testing as a clinical tool for the evaluation of impotence and provide new treatment alternatives for erectile dysfunction.
ACKNOWLEDGMENTS
We would like to thank Dr. Helmut S. Schmidt, Dr. Jian-Sheng Lin, and Dr. Jeffrey Brown for their advice and critical review of the manuscript. Research supported in part by INSERM U52, CNRS URA 1195, Claude Bernard University and the Medical College of Ohio at Toledo.
- 1. Baum MJ and Everett BJ (1992)
Increased expression of c-fos in the medial preoptic area after mating in male rats: role of afferent inputs from the medial amygdala and midbrain central tegmental field.
Neuroscience 50; 627-646. - 2. Chen KK, Chan JYH, Chang LS, Chen NIT and Chan
SHH (1992)
Elicitation of penile erection following activation of the hippocampal formation in the rat.
Neurosci Lett 141; 218-222. - 3. Fernandez-Fewell GD and Meredith M (1995)
Facilitation of mating behavior in male hamsters by LHRH and AcLHRh3-10: Interaction with the vomeronasal system.
Physiol Behav 5 7; 213-22 1. - 4. Fisher C, Gross I and Zuch J (1965)
Cycles of penile erection synchronous with dreaming (REM) sleep: Preliminary report.
Arch Gen Psychiat 12; 29-45. - 5. Gandolfo G, Glin L, Zernicki B and Gottesmann C
(1985)
The chronic "cerveau isolé" rat shows late and exceptional occurrence of paradoxical sleep.
Arch Ital Biol 126; 223-226. - 6. Giuliano FA, Rampin 0, Benoit G, and Jardin A (1995)
Neural control of penile erection.
Urologic Clinics of North America 22; 747-766. - 7. Halstead LS, Dimitrijevic M, Karacan I and et al
(1984)
Impotence in spinal cord injury: Neurophysiological assessment of diminished tumescence and its relation to supraspinal influences.
Curr. Concepts Rehab Med 1; 8-14. - 8. Hanada Y and Kawamura H (1981)
Sleep-waking electrocorticographic rhythms in chronic cerveau isolé rats.
Physiol Behav 26; 725-728. - 9.
Holmes CJ and Jones BE (1994)
Importance of cholinergic, GABAergic, serotonergic and other neurons in the medial medullary reticular formation for sleep-wake states studied by cytotoxic lesions in the cat. Neuroscience 62; 1179-1200. - 10. Holmes G, Chapple W, Leipheimer R and Sachs B
(1991)
Electromyographic analysis of male rat perineal muscles during copulation and reflexive erections.
Physiol Behav 49; 1235-1246. - 11. Jacobs BL
Overview of the activity of brain monoaminergic neurons across the sleep-wake cycle.
In Sleep: Neurotransmitters and Neuromodulators (ed. Wauquier A, Gaillard JM, Monti JM and Radulovacki M) New York Raven Press 1985; 1-14. - 12. Jones BE (1989)
Basic mechanisms of sleep-wake states.
In Principles and Practice of Sleep Medicine (ed. Kryger MH, Roth T and Demerit WC), Philadelphia: W.B. Saunders Company, 1989: 121-138. - 13. Jones BE (1991)
Paradoxical sleep and its chemical/structural substrates in the brain.
Neuroscience 40; 637-656. - 14. Jouvet
M (1962)
Recherches sur les structures nerveuses et les mecanismes responsables des différentes phases du sommiel physiologique.
Arch Ital Biol 100; 125-206.
Texte intégral en html - 15. Jouvet
M (1967)
Neurophysiology of the states of steep.
Physiol Rev 47;117-177.
Texte intégral en html - 16.
Kanamori N, Sakai K, Sei H, Bouvard A, Salvert D, Vanni-Mercier G and
Jouvet M(1995)
Effects of decerebration on blood pressure during paradoxical sleep in cats.
Brain Res Bull 37; 545-549. - 17. Karacan I, Dervent A, Salis PJ and Ware JC (1978)
Spinal cord injuries and NPT.
Sleep Res 7; 261. - 18. Karacan I, Dimitrijevic M, Lauber A, Ware JC,
Halstead L, Attia S, Altinel A and Salis P (1977)
Nocturnal penile tumescence (NPT) and steep stages in patients with spinal cord injuries.
Sleep Res 6; 52. - 19. Karacan I, Goodenough DR, Shapiro A and Starker
S (1966)
Erection cycle during sleep in relation to dream anxiety. Gen Psychiat 15; 183. - 20. Kondo Y (1992)
Lesions of the medial arnygdala produce severe impairment of copulatory behavior in sexually inexperienced male rats.
Physiol Behav 51; 939-943. - 21. Marson L, List MS and McKenna KE (1992)
Lesions of the nucleus paragigantocellularis alter ex copula penile reflexes.
Brain Res 592; 187-192. - 22. Marson L and McKenna KE (1990)
The identification of a brainstem site controlling spinal sexual reflexes in male rats.
Brain Res 515; 303-308. - 23. Marson L and McKenna KE (1992)
A role for 5-hydroxytryptamine in descending inhibition of spinal sexual reflexes.
Exp Brain Res 88; 313-320. - 24. Marson L and Mckenna KE (1994)
Stimulation of the hypothalamus initiates the urthrogenital reflex in male rats.
Brain Res 638; 103-108. - 25. Melis MR, Stancampiano R and Argiolas A (1994)
Penile erection and yawning induced by paraventricular NMDA injection in male rats are mediated by oxytocin.
Pharmacol Biochem Behav 48; 203-207. - 26. Michel F, Klein M, Jouvet D, Valatx JL and Jouvet
M (1961)
Etude polygraphique du sommeil chez le rat.
CR Acad Sci 12; 2389-2392. - 27. Nakata K and Kawamura H (1986)
ECoG sleep-waking rhythms and bodily activity in the cerveau isoIé rat.
Physiol Behav 36; 1167-1172. - 28. Paxinos G and Watson C
The Rat Brain in Stereotaxic Coordinates. Tokyo: Academic Press, 1986. - 29. Sachs BD and Bitran D (1990)
Spinal block reveals roles for brain and spinal cord in the mediation of reflexive penile erections in rats.
Brain Res 528; 99-108. - 30. Sachs BD and Garinello LD (1978)
Interaction between penile reflexes and copulation in male rats.
J Comp Physiol Psychol 92; 759-767. - 31. Sachs BD and Garinello LD (1979)
Spinal pacemaker controlling sexual reflexes in male rats.
Brain Res 171; 152-156. - 32. Sakai K and Jouvet M (1980)
Brain stem PGO-on cells projecting directly to the cat dorsal lateral geniculate nucleus. Brain Res 194; 500-505. - 33. Sakai K, Sastre JP, Kanamori N and Jouvet M
(1981)
State-specific neurons in the ponto-medullary reticular formation with special reference to the postural atonia during paradoxical sleep in the cat.
In Brain Mechanisms and Perceptual Awareness (ed. Pompeiano 0 and Marsan A), pp. 405-429. Raven Press, New York. - 34. Schmidt MH, Sakai K, Valatx JL, Wauquier A and
Jouvet M (1994)
The deficiency of penile erections during paradoxical sleep in the rat following spinal or mesencephalic brainstem transections.
Sleep Res 23; 34, abstract. - 35. Schmidt MH, Sakai K, Valatx JL, Wauquier A and
Jouvet M (1994)
Penile erections during paradoxical sleep are disrupted after spinal or mesencephalic transection.
J Sleep Res 3, Suppl 1, 462, abstract. - 36. Schmidt MH, Valatx JL, Sakai K, Debilly G and
Jouvet M (1995)
Validation of a new technique for the simultaneous recording of penile erections and micturition across behavioral states in the rat.
Sleep Res 24; 494, abstract. - 37. Schmidt MH, Valatx JL, Sakai K, Debilly G and Jouvet M (1995)
Corpus spongiosum penis pressure and perineal muscle activity during reflexive erections in the rat. Am J Physiol 269; R904-R913. - 38. Schmidt MH, Vatatx JL, Schmidt HS, Wauquier A and Jouvet M (1994)
Experimental evidence of penile erections during paradoxical sleep in the rat.
NeuroReport 5; 561-564. - 39. Shimura T, Yamamoto T and Shimokochi M (1994)
The medial pre- optic area is involved in both sexual arousal and performance in male rats: re - evaluation of neuron activity in freely moving animals.
Brain Res 640; 215-222. - 40. Siegel JM (1989)
Brainstem mechanisms generating REM sleep.
In Principles and Practice of Sleep Medicine (ed. Kryger MH, Roth T and Dement WC), pp. 104-120. W. B. Saunders Company, Philadelphia. - 41. Steriade M, Sakai K and Jouvet M (1984)
Bulbo-thalamic neurons related to thalamocortical activation processes during paradoxical sleep.
Exp Brain Res 54; 463-475. - 42. Valcourt RJ and Sachs BD (1979)
Penile reflexes and copulatory behavior in male rats following lesions in the bed nucleus of the stria terminalis.
Brain Res But 4; 131-133. - 43. Vanni-Mercier G; Sakai K, Lin JS and Jouvet M (1991)
Carbachol microinjections in the mediodorsal pontine tegmentum are unable to induce paradoxical sleep after caudal pontine and prebulbar transections in the cat.
Neurosc Lett 130; 41-45. - 44. Webster HH Friedman L and Jones BE (1986)
Modification of paradoxical sleep following transections of the reticular formation at the potomedullary junction.
Sleep 9; 1-23.