| Évolution électroencéphalographique des différents états de sommeil chez le chaton |
| J.L. Valatx, D. Jouvet et M. Jouvet Electroencephalography and Clinical Neurophysiology 17 pp : 218-233 (1964) |
L'existence de deux états EEG et comportementaux du sommeil chez le chat adulte (Dement 1958; Jouvet et al. 1959), dont les interrelations sont encore imprécises, rend particulièrement intéressante l'étude de leur apparition et de leur évolution après la naissance. Le chaton naît en effet avec un cerveau dont la maturation n'est pas encore achevée (Windle 1940). On peut ainsi, au cours de la maturation post natale, étudier un décalage chronologique possible dans l'apparition des deux états de sommeil qui coexistent ensuite chez le chat adulte. Bien que les données de la littérature sur l'activité électrique corticale des mammifères nouveau-nés étudiés dans des conditions d'expériences aigues soient maintenant très nombreuses (cf. Marty 1962 pour référence), il y a eu relativement peu de travaux consacrés à l'évolution post natale de l'EEG au cours du sommeil physiologique dans des conditions chroniques. Parmi celles-ci, quelques études portent sur le sommeil du chiot (Charles and Fuller 1956), du macaque (Caveness 1961; Robert de Ramirez de Arellano 1961), du cochon (Pampiglione 1961) mais l'existence de deux états comportementaux ou EEG du sommeil n'est pas envisagée. Chez le chaton, I'évolution polygraphique des deux phases du sommeil a été décrite dans une note préliminaire (Jouvet et al. 1961) tandis que l'activité hippocampique au cours de ces stades faisait l'objet d'autres travaux (Cadilhac et al. 1961; Cadilhac et Passouant-Fontaine 1962). Cependant, Marley et Key (1963) ne distinguent pas différents états de sommeil dans leur étude sur la maturation de l'EEG et du comportement du chaton. Le présent travail décrit l'évolution chronologique des aspects polygraphiques des deux phases du sommeil chez le chaton, de la naissance à l'âge de 3 mois.
Parmi 40 chatons opérés à la naissance, seize ont pu être étudiés et enregistrés quotidiennement jusqu'à l'âge de 3 mois. L'implantation chronique d'électrodes corticales constituées par 4 très fines vis, fixées sur la voûte crânienne de part et d'autre des sutures coronales et lambda au moyen de résine acrylique, est réalisée sous anesthésie locale à la novocaine. L'activité électromyographique (EMG) est enregistrée au moyen de fils d'or ou d'acier placés à demeure dans les muscles de la nuque (Jouvet et al. 1959). Les chatons sont remis à leur mère qui continue à les nourrir sans se soucier des électrodes. Les électrodes oculaires sont placées latéralement de part et d'autre de l'orbite osseuse; la respiration est enregistrée à l'aide d'un caoutchouc graphité entourant le thorax de l'animal et dont les extrémités sont reliées à l'appareil EEG par l'intermédiaire d'un pont de Wheatstone. L'enregistrement est pratiqué sur un appareil EEG Alvar pendant 2 à 3 h, après une tétée, dans une cage insonorisée et chauffée à 30°C.
La décérébration est pratiquée également sous anesthésie locale. Après incision de la peau et ouverture de l'os à l'aide de ciseaux fins, un leucotome est orienté manuellement en avant de la tente du cervelet et descendu jusqu'à la base du crâne. Les soins postopératoires sont importants car le chaton décérébré ne téte pas spontanément: il faut donc le gaver toutes les 3 h avec du lait à l'aide d'une sonde introduite dans la cavité buccale, évacuer la vessie par pression et le maintenir à une température constante dans une couveuse réglée à 37°C.
L'étendue des lésions a été contrôlée par des coupes sagittales du tronc, colorées selon les méthodes de Nissl et Luxol fast Blue.
Résultats
A. Chaton intact
Nous avons distingué trois périodes principales au cours des six premières semaines, en nous basant sur des critères comportementaux et électroencéphalographiques.
1. De la naissance au 6e jour (poids de 110 à 130 g à la naissance)
Trois stades comportementaux de vigilance sont observés mais l'activité EEG ne permet pas de faire de distinction entre l'éveil et les phases de sommeil. L'observation attentive de l'animal et l'enregistrement polygraphique sont nécessaires (Fig. 1).
(a) Éveil: comportement. Après la naissance et la section du cordon ombilical, le chaton reste inerte sur le flanc. Puis, au bout de 2 min apparaît la réaction posturale: le chaton se retourne et se met en décubitus ventral. Il s'approche ensuite de sa mère qui l'appelle à petits cris, en "crawlant" avec les pattes antérieures et balançant la tête de part et d'autre; 10 à 12 min suffisent pour trouver la mamelle.
Jusqu'à l'âge de 7 jours, il n'y a pas de modifications notables du comportement: tournant à droite et à gauche la tête relevée ou au ras du sol, les paupières closes, avec des miaulements fréquents, le chaton se déplace lentement et maladroitement en se traînant sur le sol à l'aide de ses pattes antérieures; les pattes postérieures sont allongées dans le prolongement du corps les premiers jours, puis se replient sur le côté en donnant une démarche "en canard". Il n'existe pas de synergie entre le train antérieur et postérieur. Le chaton réagit en sursautant à une stimulation intense sonore ou somesthésique (pincement de la peau) mais ne réagit pas à la piqûre.
Les enregistrements polygraphiques que nous avons pratiqués à partir d'une heure après la naissance, montrent un rythme cardiaque rapide aux environs de 300 pulsations par minute.
La respiration est très irrégulière, tantôt rapide, superficielle (120/min) pendant quelques secondes, tantôt plus lente (40-60/min). L'électromyogramme est constitué par une activité d'une fréquence de 50 à 60 c/sec d'une amplitude de 100 à 200 microV.
L'activité électrique corticale pariéto-occipitale a une fréquence de 8-12 c/sec de bas voltage (20 à 30 microV). Des bouffées irrégulières en fuseaux de 12-14 c/sec et de 50 à 80 microV, relativement espacées, toutes les 5 à 6 sec, au début, tendent à se rapprocher vers la fin de la première semaine sans se fusionner (Fig. 1,1).
Au niveau des dérivations frontales, l'activité électrique est plus rapide, plus régulière et de moins grande amplitude.
Lors des stimulations nociceptives (pincement de la peau), un aplatissement du tracé de très courte durée (quelques secondes) est enregistré inconstamment. Cette réaction "s'habitue" alors très rapidement, mais le plus souvent les réactions d'éveil comportemental ne s'accompagnent d'aucune variation EEG décelable.
Chez le chaton prématuré (moins de 80 g), il existe des périodes d'activité électrique nulle alternant avec des fuseaux réguliers de 7 à 10 c/sec (Fig. 5, pm).
(b) Le "sommeil calme". L'éveil est parfois suivi d'une très courte phase intermédiaire où le chaton est au repos; aucun mouvement n'est observé ni enregistré, mais il n'y a pas encore de véritable posture de sommeil. Sur le tracé EMG on observe tantôt une abolition totale, tantôt une activité musculaire minime.
L'activité EEG ne se modifie pas et reste identique à celle de l'état de veille. Ces phases de "sommeil calme" sont de très courte durée (1 à 2 min) et sont inconstantes les 2 premiers jours (Fig. 1,II). Le plus souvent en effet, I'état de veille est interrompu périodiquement et parfois soudainement par des phases de "sommeil avec secousses".
La phase de 'sommeil calme" sans activité EMG coexiste avec la phase de "sommeil calme" avec activité EMG jusqu'à la 6e semaine environ, où elle disparaît progressivement et totalement. Il ne persiste alors que la phase de sommeil calme avec activité EMG qui correspond au sommeil avec ondes lentes corticales de l'adulte (Fig. 9).
(c) Le "sommeil avec secousses". Cet état est caractéristique: après une courte phase inconstante de calme, I'animal présente des mouvements différents de ceux de l'état de veille. Ce sont des secousses rapides de la tête accompagnées de mouvements des vibrisses, des oreilles, des paupières et parfois des globes oculaires, des trémulations des membres portant sur les racines et les extrémités où l'on peut observer des mouvements rapides des doigts, enfin des vermiculations du tronc et de la queue. Certains mouvements semblent plus adaptés: grattage, succion, déglutition (dans quelques cas nous avons vu des chatons présenter cette phase pendant qu'ils tétaient leur mère). Enfin, des secousses globales du corps et des sursauts entraînent le déplacement du chaton et provoquent parfois son réveil. En somme, ce sont des mouvements rapides que le chaton est incapable d'accomplir le plus souvent spontanément à l'état de veille. A ce stade, une stimulation auditive intense ne réveille pas le chaton, pour cela une stimulation somesthésique forte (pincement) est nécessaire. Le réveil se fait dans un sursaut ou progressivement par extension de la tête et des quatre membres avec baillement.
La durée moyenne de ces phases de sommeil augmente de jour en jour: elles sont brèves les 2 premiers jours (1 à 3 min), tandis qu'à la fin du premier septenaire, elles peuvent durer de 7 à 10 min (Fig. 1, II). Leur durée moyenne est alors de 3 min.
(d) Relations entre l'éveil et les deux états de sommeil. La période de "sommeil avec secousses" peut survenir immédiatement après l'éveil, aussi bien que le "sommeil calme" avec ou sans activité EMG. Ces différents cycles comportementaux (éveil-sommeil calme-sommeil avec secousses) au cours de la première semaine, n'ont ainsi pas de rapport chronologique régulier. L'éveil représente 40%; le "sommeil avec secousses" 40 à 50%, le "sommeil calme" 10 à 20% de la durée totale des enregistrements (Fig. 8, A; 9).
Résultats
A. Chaton intact
2. De 8 à 15 jours
(Fig. 2)
Au 8e jour les paupières se décollent graduellement. Dès leur ouverture, les réflexes cornéen et photo-moteur sont obtenus.
(a) Eveil: aspect comportemental. Vers le 10e jour, le chaton commence à prendre appui sur le train antérieur et à soulever l'abdomen qui touche toujours le sol. A la fin de la 2e semaine, le déplacement est lent et encore instable, avec appui sur les quatre pattes, I'abdomen ne touche plus le sol, les mouvements des membres postérieurs sont synchronisés avec ceux des pattes antérieures. Bien qu'il ait les yeux ouverts, le chaton ne présente pas encore d'orientation visuelle ni de clignement à la menace. Les mouvements d'orientation auditive débutent pour des sons aigus. Enfin, le chaton réagit à la piqûre ou au pincement par le retrait du membre stimulé et en tournant la tête du même côté.
Aspect polygraphique : (Fig. 2, I) Le rythme cardiaque est encore rapide (230/min) à la fin de la 2e semaine. La respiration est toujours irrégulière et rapide (80/min). Les mouvements spontanés des yeux sont peu nombreux, lents et de faible amplitude. Le tracé EEG reste semblable à celui enregistré au cours de la première semaine: il est représenté par une activité électrique à 12-15 c/sec en bouffées irrégulières, d'une amplitude de 80 à 100 microV. L'activité des régions frontales a une amplitude plus réduite que les régions occipitales.
La réaction d'arrêt corticale est inconstante. Elle se caractèrise par une diminution du voltage du tracé, de courte durée (inférieure à 20 sec), sans accélération de sa fréquence. Dans les conditions d'éveil intense, après une stimulation nociceptive, on peut obtenir des bouffées de 15 à 20 c/sec de grande amplitude (100 microV).
(b) Le "sommeil calme" (Fig. 2,II). Les phases de "sommeil calme" augmentent peu à peu de durée. Le chaton, les yeux fermés, commence alors à adopter une posture de sommeil "en boule".
L'enregistrement polygraphique montre la persistance d'une activité électrique avec fuseaux de 10 à 15 c/sec, de 80 à 100 microV, d'une durée de 1 à 2 sec, survenant 8 à 10 fois par minute. Le tracé est légèrement plus lent que celui de l'état de veille et les fuseaux sont moins nombreux mais il n'y a aucune onde lente corticale aussi bien pendant la phase de sommeil calme avec EMG que sans EMG.
(c) Le "sommeil avec secousses": aspect comportemental. Au cours du second septenaire, les secousses paroxystiques et les sursauts globaux du corps diminuent. Il ne persiste que des secousses discrètes de la tête avec des mouvements des oreilles, des vibrisses, des globes oculaires, ainsi que des mouvements fins des extrémités des pattes.
Aspect polygraphique (Fig. 2, III). Contemporaine de la déhiscence palpébrale, I'activité électrique cérébrale recueillie au niveau des régions postérieures se modifie profondément: elle est alors constituée par une activité caractéristique en fuseaux de 15 à 18 c/sec, de 40 à 80 microV, survenant par bouffées de 2 à 3 sec. Ces fuseaux, d'abord irréguliers (8e-9e jour) tendent à se régulariser et à fusionner en une ac tivité continue de haut voltage (80 microV) (11e-12e jour). Ensuite cette activité devient plus microvoltée (30 microV) pendant de courtes périodes (15 à 20 sec). Au 15e jour, une activité rapide de 20 à 30 c/sec continue, de bas voltage (20 à 30 microV) s'observe pendant toute la durée de ces phases de sommeil qui peuvent atteindre 7 à 10 min, alors qu'au cours de l'éveil, la durée de l'activité rapide de bas voltage reste toujours inférieure à une minute. Le rythme cardiaque se ralentit (200/min). Les mouvements des globes oculaires contemporains des sursauts de la tête au cours de la première semaine surviennent maintenant de facon indépendante. Ils sont rapides, horizontaux et verticaux, survenant par bouffées au nombre de 6 à 8 par min (Fig. 7, B et C). La phase de "sommeil avec secousses" a une durée moyenne de 5 min et survient six à sept fois par heure. Elle est presque toujours précédée par une phase de "sommeil calme".
Le pourcentage des différents états de vigilance est le suivant: - Eveil: 30 à 35% - Sommeil calme sans EMG: 25%- Sommeil calme avec EMG: 10%- "Sommeil avec secousses": 40 à 50% (Fig. 8, B; 9).
En résumé, le phénomène caractéristique de la seconde semaine post-natale est l'apparition d'une activité rapide corticale de bas voltage pendant toute la durée du "sommeil avec secousses", alors qu'au cours de l'éveil on ne peut enregistrer que de courtes périodes, phasiques, d"'activation corticale".
Résultats
A. Chaton intact
3. De la troisième à la sixième semaine
(Fig. 3)
(a) Eveil. Au cours de la 3e semaine, le chaton parachève rapidement ses acquisitions comportementales et électroencéphalographiques.
A partir du 21e jour, le chaton commence à suivre un objet du regard, d'abord dans le sens horizontal, puis dans le sens vertical. Au 23e jour, il s'assoit sur le train postérieur, puis se gratte l'oreille avec la patte arrière (24 jours), fait sa toilette en passant sur le museau la patte antérieure préalablement lèchée (26 jours), joue, gratte la sciure (27 jours), court (29 jours), grimpe le long de la porte grillagée de la cage (30 jours). Le chaton boit seul, spontanément, dans une tasse au 32e jour, mange de la viande (34 jours), se tapit à l'affut (42 jours). Le clignement à la menace apparaît vers le 43e jour. Enfin, vers le 50e jour, le chaton "boxe", se dressant sur les pattes arrières, il tape avec les pattes avant sur les doigts que l'on présente. Ainsi, en 3 semaines, le chaton a acquis toutes les réactions comportementales essentielles qui lui permettent de vivre seul, loin de sa mère.
Le tracé électroencéphalographique de veille évolue peu à peu vers l'état adulte (Fig. 3, I; 5; 6). L'activité électrique augmente de fréquence (18 à 20 c/sec) pendant l'éveil intense. A la fin de la 5e semaine, la "réaction d'arrêt", constituée par une activité rapide (20 à 25 c/sec) de bas voltage, devient tonique: sa durée peut être alors supérieure à 2 min. Elle est alors identique à celle de l'adulte. La durée de l'état de veille par rapport au temps d'enregistrement varie peu (25%).
(b) Le "sommeil calme". A l'endormissement, le plus souvent, il apparaît une activité comportementale stéréotypée: léchage des pattes antérieures et du museau: "le chaton fait sa toilette". Puis il adopte une posture de sommeil "en boule" ou "à plat ventre", les paupières closes avec parfois des mouvements pendulaires des globes oculaires.
L'enregistrement EEG (Fig. 3,II) montre un ralentissement progressif de l'activité électrique corticale au cours de la 3e et de la 4e semaine, de 7 à 9 c/sec au 21e jour, elle diminue à 2-4 c/sec avant la fin du premier mois. Les ondes lentes sont amples de 150 à 200 microV, diffuses, irrégulières et surviennent dès l'endormissement.
Les fuseaux à 10-12 c/sec présents depuis la naissance disparaissent au cours de la 3e semaine et laissent ainsi la place aux ondes lentes. Ils ne réapparaissent à l'endormissement que vers la fin de la 6e semaine. Il existe donc ainsi une période de 3 semaines au cours de laquelle les fuseaux corticaux disparaissent.
Les ondes lentes sont enregistrées aussi bien pendant la phase de "sommeil calme" avec activité EMG que sans activité EMG. Le sommeil lent sans EMG diminue et disparait vers le 45e jour (Fig. 9). Il ne persiste alors que la phase de "sommeil calme" avec ondes lentes corticales et activité EMG des muscles de la nuque, ressemblant en tous points au "sommeil lent" décrit chez l'adulte.
(c) Le "sommeil avec secousses" (Fig. 3,II). Le comportement du chaton au cours de cette phase de sommeil devient identique à celui observé chez le chat adulte pendant la phase paradoxale ou rhombencéphalique du sommeil (P.R.S.). Le chaton roulé en boule sur le flanc ou en décubitus ventral, la tête reposant sur le sol, présente des mouvements des vibrisses, des battements des oreilles et de fins mouvements des extrémités et de la queue. A travers les paupières mi-closes, on observe des mouvements rapides des yeux, nystagmiformes, horizontaux et verticaux (Fig. 7, D).
L'activité électrique cérébrale est rapide, à 25-30 c/sec, continue, de bas voltage (20 à 30 microV), tandis que l'activité EMG des muscles de la nuque est totalement abolie. Le rythme cardiaque se ralentit le plus souvent, la respiration devient irrégulière. Cette phase, dont la durée moyenne est de 5 min, revient de facon périodique cinq fois par heure. Elle représente 40 à 20% du temps total d'enregistrement (Fig. 8, C; 9).
A 90 jours, le comportement et l'activité électrique cérébrale du jeune chat au cours des différents stades de vigilance, sont semblables à ceux d'un chat adulte.
Résultats
B. Chaton décérebré
Parmi neuf chatons décérébrés à la naissance, 3 ont eu une survie suffisante (5 jours) pour que leur rythme veille-sommeil puisse être étudié.
(a) Eveil. Dès la fin de l'intervention, on observe par moments une extension de la nuque et du train antérieur, aisément vaincue à la flexion passive. ll n'y a pas de rigidité. Mis sur le dos, le chaton a des mouvements de "pédalage" des pattes antérieures, mais ne se retourne pas en décubitus ventral. Une réaction posturale lente ne réapparaît qu'environ 24 h après l'opération. Il existe des périodes de calme et d'agitation où le chaton se déplace droit devant lui rapidement. Le "crawling approach" avec balancement de la tête de part et d'autre a disparu. L'animal réagit à la stimulation nociceptive (pincement) par un cri et par le retrait du membre et sursaut de tout le corps. Remis avec la mère, il ne cherche pas la mamelle, ne tète pas lorsque le tèton est mis dans sa gueule. Cependant il miaule fréquemment et le réflexe de déglutition est obtenu facilement si l'on place du liquide sur sa langue.
(b) Sommeil. L'observation du chaton décérébré met en évidence des phases de "sommeil avec secousses" identiques à celles du chaton intact, avec les mêmes secousses de la tête et du corps et, fait paradoxal, de nombreux mouvements spontanés de succion et de déglutition qui ne sont pas observés à l'état de veille. L'activité EMG des muscles de la nuque qui est importante pendant l'éveil est complètement abolie au cours du "sommeil avec secousses". L'activité électrique recueillie au niveau des régions frontales (en avant de la section du tronc cérébral) montre une activité en fuseaux irréguliers de 6 à 12 c/sec, de 50 microV d'amplitude, alternant avec des périodes de silence électrique. Il n'y a aucune variation EEG au cours des différents stades de vigilance. L'E.K.G. révèle une bradycardie relative au cours de l'éveil et du sommeil qui persiste 3 à 4 jours après la décérébration. Le rythme respiratoire est plus lent que chez le chaton intact, très irrégulier, avec des pauses de quelques secondes. Par rapport à la durée d'enregistrement la pourcentage du "sommeil avec secousses" est de 40% et celui d'éveil 50 à 60%. Le contrôle histologique pratiqué, a montré que la section du tronc cérébral en avant des tubercules quadrijumeaux était totale (Fig. 4).
En résumé, une section totale du tronc cérébral, au niveau du mésencéphale, n'entraine pas au cours des 5 premiers jours après la naissance, de différences appréciables du cycle comportemental veille-sommeil.
Discussion
Nature des deux phases de sommeil observées chez le chaton
A. "Sommeil avec secousses"
On sait que chez le chat adulte la phase de "sommeil activé" (Dement 1958) ou "phase paradoxale" du sommeil ou "phase rhombencéphalique du sommeil" (P.R.S.) (Jouvet et al. 1959), se traduit par une activité corticale rapide et de bas voltage, associée à un rythme thêta hippocampique et à une activité de pointes monophasiques, groupées en pseudo "fuseaux" au niveau du pont, alors qu'apparaissent des mouvements rapides des yeux, des vibrisses et une disparition totale de l'activité musculaire de la nuque. Cette phase comporte un seuil d'éveil plus élevé que la phase d'ondes lentes corticales. Les structures nerveuses responsables du déclenchement de cette phase ont été délimitées par des expériences de section et de coagulation du tronc cérébral. Elles sont situées au niveau de la formation réticulée pontique (Jouvet 1962a). Les critères suivants permettent d'assimiler la phase du "sommeil avec secousses" du chaton à la P.R.S. de l'adulte.
1. Critères anatomiques. La présence de phases de "sommeil avec secousses" chez des chatons décérébrés dont l'allure polygraphique est identique en tous points à celles observées chez le chaton normal, démontre le rôle suffisant de la partie caudale du tronc cérébral dans leur apparition. Cependant, nous n'avons pas encore pu démontrer le rôle nécessaire joué par la formation réticulée pontique chez le chaton car il n'a pas été possible d'effectuer des coagulations suffisamment précises compatibles avec une survie prolongée.
2. Critères comportementaux. La similitude des signes somatiques et végétatifs du "sommeil avec secousses" du chaton avec ceux de la P.R.S. chez l'adulte est frappante.
Les mouvements rapides des yeux ont été observés dès le premier jour post natal à travers les paupières closes. Ils sont à ce stade presque toujours contemporains des mouvements des vibrisses et des secousses de la tête et il est alors difficile de les enregistrer, soit à cause des artéfacts dûs aux mouvements de la tête 7 soit parce que la potentiel cornéorétinien a une amplitude trop faible. Dès le 8e jour par contre (Fig. 7), avant même que les yeux soient ouverts, leur aspect oculographique ressemble à celui de l'adulte. Ce problème des mouvements oculaires est particulièrement intéressant car de nombreux arguments permettent de penser que la P. R.S. est l'analogue chez l'homme du sommeil onirique (Dement et Kleitman 1957; Jouvet 1962b) et il a été montré que les mouvements rapides des yeux chez l'homme peuvent avoir un rapport étroit avec l'imagerie visuelle des rêves (Roffwarg et al. 1962). Il semble difficile cependant d'admettre qu'avant l'ouverture des yeux, les chatons puissent présenter une perceptivité visuelle. Il faut donc admettre que de tels mouvements oculaires au cours du sommeil précèdent l'entrée en fonction du système visuel. A ce sujet, on peut signaler également que les mouvements des yeux du chat adulte persistent au cours de la P.R.S. même après destruction de la rétine (Mouret et al. 1963) et peuvent encore se voir chez des animaux mésencéphaliques chroniques (Jouvet 1962a).
Les mouvements des vibrisses et les petits mouvements des doigts, des oreilles, de la queue, sont similaires à ceux observés chez l'adulte. Cependant, il est rare d'observer des secousses globales du corps chez l'adulte normal. Par contre, des secousses de tout le corps, parfois violentes, s'observent au cours des P.R.S. chez des chats adultes décortiqués ou pontiques chroniques (Jouvet 1962a). Le mécanisme de tels soubresauts musculaires reste encore inconnu: il faudrait admettre que chez le chaton comme chez l'adulte décérébré, des influences motrices descendantes extrapyramidales puissent forcer temporairement l'inhibition qu'exerce la formation réticulée inhibitrice au niveau spinal.
Tonus musculaire: sa disparition constitue chez le chat adulte l'index comportemental absolu de la P.R.S. (Jouvet et al. 1959). Chez le chaton de moins d'un mois, cet index est moins fidèle, car si la disparition de toute activité musculaire est constante au cours de chaque P.R.S. (mis à part les bouffées d'activité phasique apparaissant lors des secousses globales), par contre, il n'est pas rare de l'observer au cours de l'autre phase du sommeil (voir plus bas).
Variations végétatives: elles sont constantes et beaucoup plus nettes que chez l'adulte. La bradycardie est remarquable et s'observe aussi bien chez le chaton décérébré que normal (Fig. 9). Contrairement au chat adulte, on n'a pas observé de tachycardie au cours de cette phase. L'accélération et l'irrégularité respiratoires sont également beaucoup plus nettes que chez l'adulte.
Ces variations végétatives sont tellement évidentes que souvent on peut juger de l'état de sommeil du chaton uniquement sur leur caractère polygraphique.
A tous ces critères nous pouvons ajouter celui de la "profondeur" du sommeil, car bien que le "sommeil avec secousses" ait les apparences comportementales d'un sommeil léger, il est très difficile d'éveiller un chaton au cours de ce stade, alors que des stimulations plus faibles l'éveillent au cours du "sommeil calme". Là aussi, comme chez l'adulte, ce "sommeil avec secousses" apparaît bien paradoxal.
3. Critères EEG (Fig. 5, 6 et 9). Ils doivent être considérés selon l'age du chaton.
La première semaine en effet, I'activité corticale ne varie pas au cours des différents stades comportementaux de vigilance. La présence d'une activité rythmique ayant une allure de fuseaux irréguliers de 10 à 12 c/sec alternant avec une activité très microvoltée, est très caractéristique et a été retrouvée chez tous nos animaux, y compris les prématurés (Fig. 5). Bien que de tels éléments n'aient pas été décrits chez le chaton nouveau-né dans des conditions d'expérimentation aiguë (curarisation ou Nembutal) (Grossman 1955; Libet et al. 1941), d'autres auteurs par contre les ont retrouvés dans des conditions chroniques (Marley et Key 1963), associés à une activité de 1 à 3 c/sec de très bas voltage que nous n'avons pas observée. On sait qu'à la naissance, seules les synapses axono-dendritiques des régions superficielles du cortex sont bien développées, identiques en tous points à celles du chat adulte, tandis que les cellules nerveuses ne possèdent pas d'articulations synaptiques. Le mécanisme d'apparition de ces fuseaux pourrait être en relation avec l'activité de ces synapses (Purpura 1961).
L'absence de réactivité EEG au cours de la première semaine lors des différents stades de vigilance, soulève également de nombreux problèmes: il semble que l'on puisse l'expliquer par l'état d"'immaturation" du cortex. Celui-ci répond en effet d'une manière univoque par des ondes rythmiques régulières de 10 à 14 c/sec, similaires aux fuseaux, à toute stimulation du thalamus ou de la formation réticulée, aussi bien à des fréquences lentes qu'à des fréquences rapides qui provoquent une "activation" chez l'adulte (Purpura et al. 1960). Ainsi, au cours de la première semaine, I'activité corticale est incapable d'être modifiée par les influences ascendantes d'origine sous-corticale au cours de l'éveil et du "sommeil avec secousses", alors que les influences descendantes permettent déjà une distinction nette entre les deux états au point de vue comportemental.
Vers le 10e jour, apparaît une différenciation EEG seulement au cours du "sommeil avec secousses" (Fig. 5 et 6), caractérisée par une activité électrique rapide à 20 c/sec de grande amplitude, puis bientôt de bas voltage. C'est également à cet âge qu'a été notée (Cadilhac et al. 1961; Cadilhac et Passouant-Fontaine 1962) au cours de cette phase de sommeil, I'apparition, au niveau de l'hippocampe dorsal, d'une activité thêta régulière qui contraste avec l'activité irrégulière de l'éveil. Pour hypothétique que soit toute tentative de corrélation entre l'activité électrique corticale et le degré d'organisation neuronique et gliale du cortex, il est intéressant de noter que l'apparition d'une activité EEG rapide au cours du sommeil est contemporaine du développement des dendrites basilaires des cellules pyramidales et des cellules étoilées profondes, ainsi que des synapses axono-somatiques (Purpura 1961; Marty 1962). Il se pourrait ainsi que ces structures aient quelques rapports avec l'activité rapide corticale de la P.R.S. Par contre, I'activation corticale d'éveil n'est pas encore nette et durable au cours de la 2e semaine puisqu'on observe seulement de courtes phases (inférieures à 1 min) d'activité rapide de bas voltage lors des stimulations sensorielles les plus fortes, I'activité d'éveil EEG étant généralement constituée par un rythme thêta à 6-9 c/sec, associée à des fuseaux (Fig. 5 et 6).
Ces faits permettent ainsi de différencier chronologiquement les structures sous-corticales ou corticales responsables de l'activité rapide corticale de la P.R.S. de celles responsables de "I'activation" corticale d'éveil: système réticulé activateur ascendant (Moruzzi et Magoun 1949).
Une telle différenciation avait déjà été notée chez le chat adulte chez lequel des lésions du tronc permettent de supprimer électivement, bien que transitoirement, I'une ou l'autre de ces activités (Jouvet 1962a). Le rapport chronologique étroit qui existe entre l'apparition post natale d'une activité thêta au niveau de l'hippocampe et "I'activation" neocorticale au cours de la P.R.S. (Cadilhac et al. 1961) permet de supposer l'intervention du système limbique selon des modalités qu'il reste à préciser.
Discussion
Nature des deux phases de sommeil observées chez le chaton
B. Le sommeil calme
1. Son aspect comportemental permet aisément de l'assimiler à la phase de sommeil lent de l'adulte: absence de mouvements spontanés, respiration calme et régulière. Certaines caractéristiques sont cependant intéressantes à discuter.
Activité électrique des muscles de la nuque: chez l'adulte, on le sait, il persiste toujours une activité EMG de la nuque au cours du sommeil lent. Par contre, chez le chaton, jusqu'à la 6e semaine, cette activité est tantôt conservée, tantôt abolie pendant cette phase de sommeil. Il nous a semblé cependant que les phases de sommeil calme sans activité EMG étaient en rapport avec la posture de sommeil de l'animal. Au cours des premières semaines en effet, le chaton dort dans n'importe quelle position, en décubitus ventral, latéral ou dorsal. Dans les conditions normales, il se trouve auprès de sa mère ou entouré de ses congénères qui le réchauffent. Nos conditions d'enregistrement dans une ambiance chaude à 30°C permettent également toutes ces positions. C'est dans la posture en décubitus dorsal que s'observe le plus souvent l'abolition de l'EMG des muscles de la nuque qui se trouve ainsi relâchée au maximum. Progressivement, le chaton va adopter la position de décubitus ventral, la tête entre les pattes de devant, ou bien si la température ambiante est basse, la position "en boule" caractéristique du chat adulte. Dans cette posture, il persiste presque toujours une activité EMG de la nuque pendant le sommeil calme. Il semble ainsi que cette posture de sommeil lent soit acquise au cours de développement ontogénétique et qu'elle puisse avoir quelques rapports avec les mécanismes de lutte contre le froid.
2. Activité EEG: Alors que pendant les 3 premières semaines l'activité EEG de sommeil calme est presque identique à celle de l'éveil (fuseaux de 12 à 14 c/sec), vers la 4e semaine les fuseaux disparaissent et sont remplacés par des ondes lentes de 1 à 4 c/sec de haut voltage, souvent plus lentes et plus amples que chez l'adulte (Fig. 5). Ces fuseaux ne réapparaissent ensuite à l'endormissement que vers la 6e semaine et présentent alors l'aspect classique à 11-18 c/sec décrit chez le chat adulte (Rheinberger et Jasper 1937; Hess et al. 1953). S'il est diflicile d'expliquer la diminution ou la disparition transitoire des fuseaux de sommeil entre la 3e et la 6e semaine, I'évolution de l'EEG au cours de sommeil calme permet cependant de différencier chronologiquement l'activité de fuseaux observée à la naissance de l'activité lente qui semble coincider avec l'achèvement de la maturation corticale. C'est en effet vers la 4e semaine que les potentiels évoqués visuels ou somesthésiques corticaux acquièrent un aspect définitif identique à ceux de l'adulte (Marty 1962) et c'est à cet âge quele chaton acquiert un réflexe d'orientation visuelle. Les ondes lentes apparaissent ainsi comme l'expression d'une activité corticale hautement différenciée, et pour certains (Marty 1962) la prolifération des dendrites basilaires des cellules pyramidales à la finde la 3e semaine, jouerait un rôle important dans l'achèvement de cette différenciation. Ainsi les données acquises au cours de l'ontogenèse rejoignent les résultats d'expériences faites chez le chat adulte, qui démontrent l'origine spécifiquement corticale des ondes lentes. Il avait déjà été noté en effet ( Jouvet et Michel 1958; Sergio et Longo 1960) que les ondes lentes disparaissent définitivement après ablation totale du néocortex, tandis que l'ablation totale du thalamus (Naquet et al. 1964) laisse persister des ondes lentes corticales tout en supprimant totalement les fuseaux au cours du sommeil lent.
Interrelations entre les deux états de sommeil
Deux données principales apparaissent en considérant l'évolution post-natale des deux états de sommeil (Fig. 8 et 9).
A. A la naissance, le "sommeil avec secousses" ou P.R.S. peut interrompre périodiquement l'état de veille sans phase de sommeil calme intermédiaire. Ce fait révèle d'une part que les structures nerveuses pontiques déclenchant les phénomènes comportementaux toniques et phasiques de la P.R.S. (atonie musculaire, secousses, mouvements des yeux, etc.) entrent en jeu plus précocement au cours du développement que les structures responsables du comportement de "sommeil calme"; ontogénétiquement la P.R.S. apparaît ainsi comme un archéo-sommeil (Jouvet 1962a). Il ne semble cependant pas en être ainsi au point de vue phylogénétique, car cet état de sommeil n'existe que de façon rudimentaire chez certains oiseaux (pigeon, poule), et est absent chez certains reptiles (tortue) (Klein et al. 1964). D'autre part, on doit admettre que la P.R.S. est un état de sommeil qualitativement différent du sommeil calme puisqu'il peut apparaître en l'absence de celui-ci; un tel phénomène peut s'observer également chez des chats adultes pontiques chroniques (Jouvet 1962a). Le sommeil calme va prendre par contre une importance relative de plus en plus grande au cours du développement post-natal. S'il ne constitue en effet que 5% environ de la durée du sommeil comportemental à la naissance, il occupe 60 à 70% de celui-ci à partir du 2e mois. Il devient alors une condition nécessaire à l'apparition des P.R.S. puisqu'il existe toujours une phase de sommeil lent entre l'éveil et la P.R.S.
B. Il est également intéressant de remarquer que la durée moyenne des P.R.S. (4 à 5 min) à la fin de la première semaine est à peu près identique à celle de l'adulte (6 min). C'est donc par une périodicité plus élevée et non par une durée plus grande, que s'exprime l'importance relative de la P.R.S. chez le chaton. Il est difficile cependant de reconnaître quels sont les facteurs spécifiques qui ralentissent la périodicité des P.R.S. au cours du développement: états de veille et de sommeil calme plus longs, prises de nourriture plus espacées, etc.
Ainsi les données de l'ontogenèse semblent bien révéler qu'à côté du sommeil classique (le sommeil lent), on doit considérer l'évolution d'un état particulier et spécifique du fonctionnement nerveux qui s'exprime par le sommeil paradoxal. S'il est encore d'usage de faire rentrer cet état parmi les "phases" du sommeil malgré sa précession ontogénétique et ses caractères EEG et comportementaux particuliers, rien ne permet, en fait, de savoir si cet état, qui semble être l'expression de l'activité onirique chez l'homme, n'est pas l'expression d'une fonction différente de celle du sommeil lent.
Résumé
1. L'évolution des activités corticale et musculaire au cours de l'éveil et des différents états de sommeil, a été étudiée chez des chatons porteurs d'électrodes chroniquement implantées au niveau du cortex et des muscles de la nuque.
2. Cette évolution permet de reconnaitre trois périodes au point de vue EEG et comportemental:
(a) Au cours de la première semaine postnatale, I'activité corticale ne varie pas lors des différents états de vigilance. Cependant, l'observation des animaux et l'enregistrement polygraphique permettent de reconnaître en plus de l'état d'éveil deux comportements bien distincts de sommeil: le premier appelé "sommeil avec secousses" est caractérisé par une disparition totale de l'activité EMG de la nuque, un ralentissement du rythme cardiaque, des secousses globales du corps ou des extrémités, ainsi que des mouvements rapides des yeux. Cette phase qui peut survenir immédiatement après l'éveil, constitue 40 à 50% d'une période d'enregistrement et environ 90% du sommeil comportemental. L'autre phase de sommeil appelée "sommeil calme" est inconstante. Elle ne s'accompagne d'aucune secousse musculaire.
(b) Au cours de la 2e semaine, des différenciations EEG commencent à apparaître. Une activité rapide corticale persiste au cours du "sommeil avec secousses" alors que de très brèves activations phasiques s'observent au cours de l'éveil. Au cours du "sommeil calme" il persiste une activité de fuseaux corticaux et l'activité EMG de la nuque dépend alors de la position de l'animal.
(c) Après le 20e jour, les deux phases de sommeil ressemblent alors à celles des chats adultes: la transition se fait insensiblement entre la phase de "sommeil avec secousses" et la phase paradoxale de sommeil du chat adulte, tandis que la phase de sommeil calme évolue vers la phase de sommeil lent avec conservation de l'activité EMG de la nuque et apparition d'ondes lentes de haut voltage.
3. Des données établies sur le chaton décérébré à la naissance, révèlent que la partie caudale du tronc suffit à l'apparition du "sommeil avec secousses".
4. Ces données sont confrontées aux résultats acquis chez le chat adulte. Elles révèlent que le sommeil paradoxal doit être distingué comme un phénomène qualitativement différent et ontogénétiquement plus ancien que l'état de sommeil lent. D'autre part, il apparaît que les structures responsables de l'activité rapide pendant le sommeil paradoxal entrent en jeu plus précocement au cours de l'évolution post-natale que les structures responsables de l'activation corticale d'éveil. Enfin, I'activité électrique lente et de haut voltage au cours du sommeil lent, semble être contemporaine de l'achèvement de la maturation corticale.
Summary
EEG evolution of the different states of sleep in the kitten
1 . The evolution of cortical and muscular electrical activity during waking and the different states of sleep have been studied in kittens bearing electrodes chronically implanted in the cortex and in the neck muscles.
2. Three main periods can be distinguished for the EEG and behavior during this evolution:
(a) During the first week after birth the cortical activity does not change with the various states of vigilance. Nevertheless, observation of the behavior and polygraphic recordings makes it possible to differentiate two states of sleep: the first one is called "sleep with jerks" and is characterized by total disappearance of the EMG of the neck, slowing of the heart rate, jerks of the body and of the legs, and rapid eye movements. This state may follow directly the waking state and constitutes 40 to 50% of a recording session and about 90% of total behavioral sleep. The second is called "quiet sleep" and is inconstantly present. It has no muscular jerks.
(b) During the second week some EEG variation begins to appear. A fast cortical activity persists during the period of "sleep with jerks" while only very short phasic activation patterns are seen during behavioral arousal. During "quiet sleep" there are cortical spindles and the activity of the neck muscles seems to depend largely upon the sleep posture of the kitten.
(c) From the beginning of the third week onward, the two states of sleep are similar to those of the adult cat: there is a transition from "sleep with jerks" to paradoxical sleep and from "quiet sleep" to slow sleep with high voltage slow waves.
3. Studies of kittens decerebrated at birth show that the caudal part of the brain-stem is sufficient for the appearance of "sleep with jerks".
4. These results are discussed, together with the experimental results obtained in adult cats. They show that paradoxical sleep is qualitatively different from, and ontogenetically older than, slow sleep. Furthermore, it is probable that the neural structures which are responsible for the fast cortical activity during paradoxical sleep come into play sooner after birth than the neural structures responsible for cortical arousal during waking. Finally, the onset of the high voltage slow waves of slow sleep appears to be contemporaneous with the achievement of cortical maturation.
Figure 1 : 5e jour: Tracés polygraphiques des différents états de vigilance
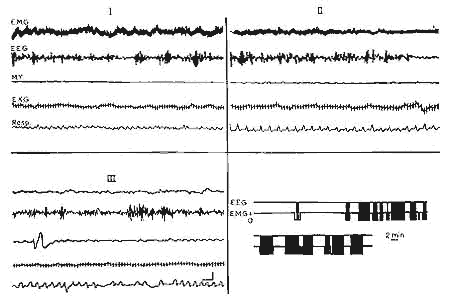
5e jour : Tracés polygraphiques des différents états
de vigilance.
Eveil (I);
Sommeil calme (II);
Sommeil avec secousses (III): Il n'y a pas de différence EEG entre
les trois stades. Représentation schématique (en bas, à
droite) des variations de vigilance au cours d'un enregistrement d'une
durce de 105 min.En noir, le sommeil avec secousses, en blanc, I'éveil.
EMG: Activité électro-myographique des muscles de la nuque;
EEG: Activité électrique frontale (bipolaire);
M.Y.: Mouvements des yeux.
Echelle pour tous les tracés EEG: 1 sec, 50 microV.
Echelle pour le schéma: 2 min.
Figure 2 : 11e jour
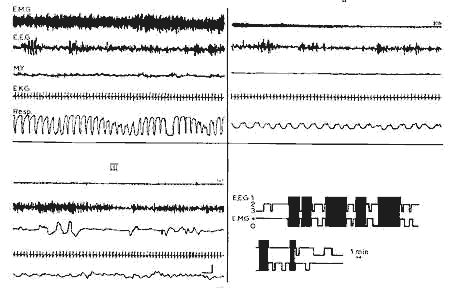
11e jour : Il n'y a pas de variation EEG entre l'éveil
(I) et le sommeil calmc(II).
Activité rapide(15 18 c/sec) en bouffées et mouvements rapides
des yeux pendant le sommeil avec secousses (III).
En bas, à droite: Représentation schématique des
trois stades de vigilance au cours d'un enregistrement de 90 min. Noter
les phases de sommeil calme sans EMG.
En noir, sommeil avec secousses; en blanc, sommeil calme; la ligne de
base représente l'éveil.
(Echelle: 1 min.)
Figure 3 : 21e jour
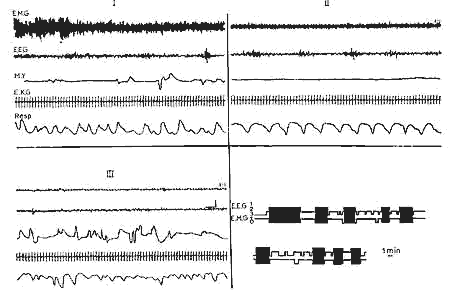
21e jour : Activité rapide de bas voltage pendant le sommeil
avec secousses (III) avec M.R.Y. semblables à ceux de l'adulte,
tandis que pendant l'éveil (I) il persiste encore quelques fuseaux.
En bas, à droite: Périodicité des différents
états pendant 105 min.
Même légende que Fig. 2. (Echelle:
1 min).
Figure 4 : Chaton décérébré
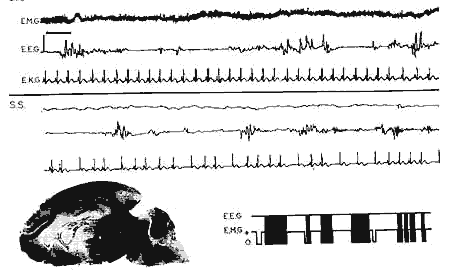
Chaton décérébré : age 6 jours. Il
n'y a pas de variation EEG pendant l'éveil et le sommeil avec secousses.
Abolition de l'EMG de la nuque et bradycardie importante pendant le sommeil
avec secousses (S.S.).
En bas, à gauche: Coupe sagittale du cerveau; la section du tronc
cérébral est totale en avant des tubercules quadrijumeaux
supérieurs.
En bas, à droite: Périodicité des phases de sommeil
avec secousses pendant un enregistrement de 60 min.
Mêmes légende et échelle que Fig.
2.
Figure 5 : Evolution de l'activité EEG au cours des trois stades de vigilance
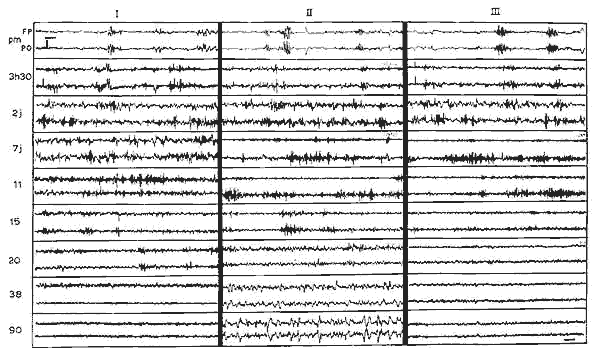
Evolution de l'activité EEG au cours des trois stades de vigilance
:
Eveil (I);
Sommeil calme (II);
Sommeil avec secousses (III) de la naissance à 90 jours.
Pour chaque âge, la première ligne représente l'activité
cérébrale enregistrée au niveau de la suture coronale,
la deuxième ligne représente l'activité occipitale.
p.m.: chaton prématuré (poids 50 g).
Les premiers jours, il n'y a pas de variation EEG au cours des trois états.
A partir du 7e jour: activité rapide (15-18 c/sec) de rapide continue
de bas voltage pendant le sommeil haut voltage pendant le sommeil avec
secousses;
A partir du 15e jour: activité avec secousses; A partir du 20e
jour: activité à 5-6 c/sec pendant le sommeil calme, qui
se ralentit progressivement. Disparition des fuseaux au cours du sommeil
lent et début d'activation corticale tonique au cours de l'éveil.
Figure 6 : Fréquences du corticogramme
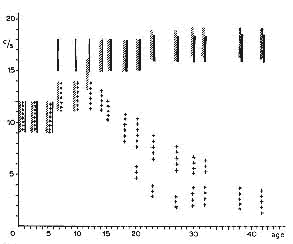
Représentation schématique de l'évolution des fréquences
du corticogramme au cours des trois états:
Eveil (hachures obliques);
Sommeil calme (croix);
Sommeil avec secousses (traits noirs verticaux) en fonction de l'âge
en jours.
Ordonnée: fréquence; abcisse: âge en jours.
Figure 7 : Evolution des mouvements des yeux
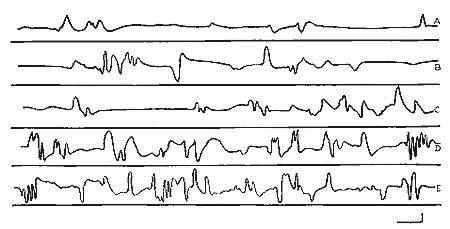
Evolution des mouvements des yeux pendant le "sommeil avec secousses, en fonction de l'âge:
A: 3 jours;, B: 7 jours; (avant la déhiscence palpébrale), C: 11 jours;, D: 15 jours;, E: Adulte.
Echelie du tracé: 1 sec, 50microV.
Dérivation bipolaire enregistrée de part et d'autre de l'orbite osseuse.
Figure 8 : Evolution de la périodicité du sommeil avec secousses

Diagramme représentant l'évolution de la périodicité
du sommeil avec secousses. Chaque cadran représente le nombre de
phases de sommeil avec secousses qui peuvent survenir dans une heure.
Les aires noires représentent la durée moyenne de ces phases
de sommeil, les aires blanches l'intervalle moyen entre les phases de
sommeil
A: 1ère semaine; durée moyenne S.S.: 4' ; intervalle moyen:4'40''
B: 2e semaine; durée moyenne S.S.: 4'44''; intervalle moyen: 4'
C: 1er mois ; durée moyenne S.S.: 5' ; intervalle moyen: 7'08"
D: 2e mois ; durée moyenne S.S.: 7'47"; intervalle moyen: 8'12"
E: 3e mois ; durée moyenne S.S.: 4'55"; intervalle moyen: 11'05"
F: Adulte ; duréemoyenne S.S.:6'20"; intervallemoyen: 23'40"
Figure 9 : Evolution post-natale du pourcentage des états de vigilance

Diagramme schématisant l'évolution post-natale du pourcentage
des états de vigilance au cours d'une durée d'enregistrement
de 3 h. En pointillés: Eveil; Flachures verticales: "Sommeil calme"
avec activité EMG de la nuque; Croisillons: "Sommeil calme" sans
activité EMG de la nuque, Hachures obliques: "Sommeil avec secousses".
La limite entre le fond continu et les bandes blanches verticales représente
l'âge à partir duquel l'aspect de l'activité cérébrale
devient semblable à celui de l'adulte pour chaque état.
Abcisse: pourcentage relatif de chaque état pour un enregistrement
de 3 h.
Ordonnée: âge en jours.