| La génétique du sommeil et du rêve |
| Jean-Louis Valatx La Recherche 1976 |
Chacun sait qu'il existe des différences notables entre individus en ce qui concerne la durée du sommeil quotidien: six heures de sommeil ou même moins suffisent à certains pour juger qu'ils ont passé une bonne nuit; de "gros dormeurs" estiment au contraire ne pas avoir suffisamment dormi s'ils n'ont pas eu dix heures ou plus de sommeil. Actuellement des recherches sur le sommeil de la souris montrent que le caractère "gros dormeur" ou "petit dormeur" dépend vraisemblablement de l'hérédité, de même que la durée du rêve au cours du sommeil.
La durée des phases du sommeil
Le sommeil est constitué de la succession périodique de deux états : le sommeil calme au cours duquel l'activité électrique cérébrale se manifeste par des ondes lentes sur l'électroencéphalogramme; le sommeil paradoxal, au cours duquel survient le rêve est caractérisé par l'activité électrique cérébrale s emblable à celle de l'éveil, tandis que le corps du dormeur est paralysé, sourd et aveugle. En outre, on peut observer des mouvements des globes oculaires, des mouvements des extrémités des membres, des variations cardiorespiratoires et une activité électrique cérébrale très particulière: l'activité phasique ponto géniculo-occipitale qui apparaît en même temps que les mouvements. Les structures nerveuses et les mécanismes biochimiques du déclenchement et de l'entretien de ces deux états commencent à être connus. La survenue et l'alternance sommeil lent sommeil paradoxal n'ont été observées que chez les oiseaux et les mammifères. Chez ces derniers, la durée de chaque type de sommeil est très variable selon les espèces. De même chez l'homme, la durée peut varier de six à douze heures selon les individus. Ces différences inter-spécifiques ou inter-individuelles sont-elles dues à l'environnement ou à des facteurs héréditaires transmis selon les lois de Mendel ? Comme pour tous les phénomènes biologiques, la réponse doit être intermédiaire : le comportement de sommeil serait la résultante de l'interaction du génome et du milieu. En effet, si on connaît bien les facteurs du milieu qui perturbent le sommeil: température ambiante, rythme lumière-obscurité, alimentation, etc., ainsi que certains facteurs psychologiques (anxiété, émotion, etc.), les facteurs héréditaires influencant le sommeil sont peu connus. Récemment, une équipe du laboratoire de médecine expérimentale (Unité 52 de l'lnserm) comprenant R. Cespuglio, G. Rondot, G. Faradji, L. Paut, M. Touret et J.L. Valatx, à l'université Claude Bernard de Lyon, a pu mettre ces facteurs en évidence, permettant d'établir un modèle expérimental simple pour approfondir nos connaissances sur les mécanismes et les fonctions du sommeil.(2-3)
Le sommeil des hybrides
Cette étude génétique du sommeil ne peut être réalisée que chez un mammifère (qui montre une alternance de sommeil lent et paradoxal) dont la carte chromosomique est déjà bien connue. Pour le moment, la souris est l'animal de choix avec ses quelque trois cents lignées pures et ses cent cinquante mutants neurologiques (fig. 1). Une lignée est dite pure après vingt générations de croisements consanguins. Tous les individus d'une même lignée ou souche sont tous identiques (comme des jumeaux monozygotes) et réagissent de la même manière à une situation expérimentale donnée. (3) Grâce à l'enregistrement polygraphique classique de l'activité électrique cérébrale, du tonus musculaire et des mouvements des yeux, nous avons étudié le rythme veille-sommeil de plusieurs souches de souris, des hybrides de 1ère génération (F1) entre les souches les plus différentes, et des croisements en retour (F1 x parents). On a pris soin que les conditions expérimentales soient rigoureusement semblables pour toutes les souris (température constante, rythme fixe lumière-obscurité de douze heures, alimentation identique). Les expériences proprement dites ont consisté à enregistrer simultanément et en continu pendant une semaine, le sommeil de cinq animaux de souches différentes. Les résultats mettent en évidence que chaque souche de souris est très homogène et présente des caractéristiques propres de durée de sommeil et d'organisation temporelle des états de sommeil (fig. 2). Le sommeil des hybrides diffère selon les croisements. Par exemple, les hybrides, résultant du croisement d'une souris de la lignée CBA avec une souris de la lignée BR, ressemblent en tous points aux parents CBA, tandis que les hybrides C x BR présentent une recombinaison des différents paramètres: durée du sommeil lent du parent C, durée du sommeil paradoxal du parent BR et rythme circadien du parent C. Enfin, la durée du sommeil des hybrides C x B6 est intermédiaire, avec le rythme circadien de C. Les animaux issus des croisements en retour (F1 x parents) sont tous différents étant donné la recombinaison des : paramètres. Cependant pour chaque paramètre, les animaux peuvent être classés en trois groupes: ceux qui ressemblent aux parents et les intermédiaires. (4)
Ces résultats indiquent:
- que chaque état de sommeil (sommeil lent, sommeil paradoxal) a un déterminisme indépendant de l'autre;
- que plusieurs gènes sont nécessaires pour déterminer un état de sommeil avec un effet de gène majeur comme dans le croisement CBA x BR;
- que non seulement la durée journalière du sommeil mais aussi sa répartition au cours de la journée est influencée par des facteurs héréditaires.

Figure 1 Enregistrement des activités durant le cycle veille-sommeil. On a adapté à cette souris un connecteur miniature fixé au crâne par du ciment dentaire, où sont soudées dix électrodes destinées à capter non seulement l'activité électrique cérébrale, mais aussi l'activité électrique liée au tonus musculaire et à celle du mouvement des yeux. Les électrodes sont reliées à l'enregistreur par un cable souple de sorte que l'animal peut bouger librement. |
La structure des groupes de neurones
Pour aller plus loin dans cette "dissection génétique" du comportement de sommeil, nous avons cherché à mettre en évidence une liaison des gènes influencant le sommeil avec des gènes dits "marqueurs" déjà connus et localisés sur la carte chromosomique de la souris. Nous avons choisi un système pluri-génique: le système d'histocompatibilité qui gouverne les antigènes de surface des cellules (système H-2), équivalent du système HLA de l'homme. Nous avons donc étudié les sept souches recombinantes obtenues par Bailey. Chacune de ces souches, issues de la génération F2 du croisement entre les souris B6 et C, représente une recombinaison (stable et différente de la souche voisine) des allèles B6 et C de tous les gènes d'histocompatibilité. Pour chacun des gènes H, la distribution des allèles B6 et C dans les sept souches est connue. Notre étude consistait à caractériser le sommeil de ces souches recombinantes: ressemblait-il à celui de l'un des deux parents originels (B6 ou C) ? Nos résultats indiquent que, pour la durée et le rythme de sommeil paradoxal, la distribution des allèles B6 et C ressemble à celle de certains gènes H (HW19, HW22). (6) Si nos résultats ne sont pas des artefacts, c'est-à-dire, si les gènes influencant le sommeil sont liés directement ou indirectement aux gènes du système d'histocompatibilité, il est possible d'émettre l'hypothèse suivante: ces gènes déterminant les antigènes de surface des cellules, les différences observées entre les souris B6 et C sont, peut-être, la conséquence de modifications de la structure des groupes de neurones (nombre de récepteurs, de neurones) impliqués dans la régulation du sommeil. Actuellement, nous essayons de vérifier cette hypothèse par l'étude anatomique comparée (souris B6 et C) des groupes cellulaires responsables du déclenchement et de l'entretien des deux états de sommeil.
Des schémas moteurs programmés
Les facteurs héréditaires n'interviennent pas seulement dans le déterminisme de la durée et du rythme de sommeil. En effet, à la suite de l'étude systématique des mouvements oculaires du sommeil paradoxal, nous avons eu la surprise de constater que leur nombre et leur répartition dans le temps (mouvements isolés ou groupés) sont identiques chez tous les animaux d'une même souche et différents d'une souche à l'autre (fig. 3). (2) Chez les hybrides C x BR dont la durée du sommeil paradoxal ressemble à celle du parent BR, les mouvements des yeux ne sont pas différents de ceux du parent C. Ainsi, les activités, phasiques des neurones du tronc cérébral à l'origine des mouvements des yeux au cours du sommeil paradoxal ne sont pas aléatoires et pourraient donc refléter un codage génétique de certaines informations. Ces résultats surprenants associés à ceux obtenus chez le chat à la suite de certaines lésion du tronc cérébral (produisant des animaux qui vivent leurs rêves) sont un élément en faveur de la théorie sur la programmation au cours du sommeil paradoxal des schémas moteurs typiques de l'espèce et leur modification sous l'influence de l'environnement.(7). Le sommeil paradoxal pourrait être comparé à un tableau dont le cadre et le support seraient représentés par sa durée et le contenu par les activités phasiques (mouvements oculaires). Les deux éléments (cadre, contenu) paraissent avoir un déterminisme indépendant: les souris BR et B6 ont la même durée de sommeil paradoxal mais les mouvements des yeux sont différents. A la limite, il doit être possible d'obtenir des souris mutantes avec seulement le cadre, c'est-à-dire, avec du sommeil paradoxal sans activité phasiques. Ces animaux seraient un précieux modèle pour la compréhension de la génèse des activités phasiques, base indispensable pour élucider les fonctions du sommeil paradoxal.
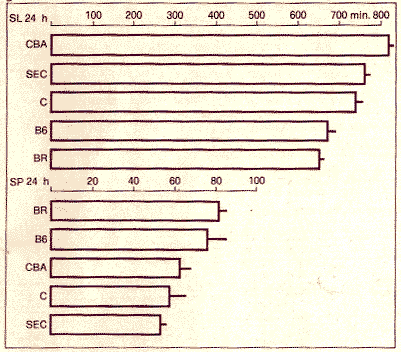
Figure 2. Durée du sommeil lent (SL) et du sommeil paradoxal (SP) par vingt-quatre heures chez cinq lignées pures de souris. On notera que les souris B6 et BR assoclent la plus courte durée de sommell lent avec la plus longue durée de sommeil paradoxal.
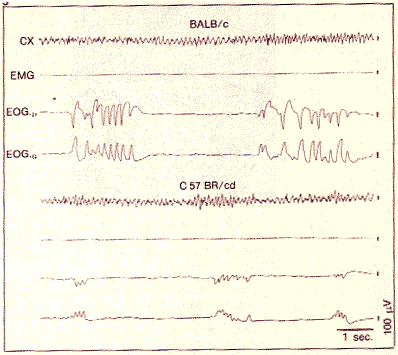
Figure 3. Enregistrement polygraphique du sommeil paradoxal des
souris BR (C57BR) et C (BALBc). CX: actité cérébrale,
EMG: tonus musculalre
EOG: mouvements des yeux.
On notera que les mouvements des yeux de BR surviennent par petits groupes alors que ceux de C forment des groupes plus importants.