| Animales rêveries |
| Mehdi Tafti , Science et Avenir Hors-Série Le Rêve Dec. 96 |
 |
Par Mehdi Tafti, |
CAFARD, DAUPHIN, TATOU, ELEPHANT...
Si les critères électrophysiologiques qui définissent les deux états de sommeil - lent et paradoxal - chez l'homme sont en général applicables aux mammifères et aux oiseaux, il n'en va pas de même pour les poissons ou les reptiles. L'absence de critères physiologiques standardisés pour ces derniers limite notre compréhension du sommeil en général, du sommeil paradoxal en particulier, et ceci pour la grande majorité des espèces vivantes.
Le premier des signes comportementaux d'un état de sommeil est probablement l'absence de mouvements, ou plutôt la présence d'un rythme d'activité-repos. Toutefois, la quiescence ne doit pas être confondue avec le sommeil, car de nombreux animaux supérieurs passent une grande partie de leur temps dans un état dit de veille calme. A l'absence ou à la diminution d'activité doivent s'ajouter d'autres critères comme une diminution de réponse aux stimuli externes ou une posture stéréotypée. Enfin, cet état doit être facilement réversible pour être distingué des états de coma, d'hypothermie ou d'hibernation.
Ces critères, s'ils nous aident à définir le sommeil en général, ne sont pas encore suffisants pour être appliqués à toutes les espèces. Nous avons encore plus de mal à définir le sommeil paradoxal par des indices autres que neurophysiologiques. Néanmoins, les corrélâts comportementaux du sommeil paradoxal sont l'atonie posturale, les mouvements oculaires et les brusques mouvements des extrémités.
A l'aide essentiellement de ces critères comportementaux, un état proche du sommeil peut être défini chez les insectes, les amphibiens et les poissons. Ils présentent un rythme d'activité-repos qu'ils peuvent maintenir même s'ils sont privés de l'alternance lumière-obscurité, comme par exemple lorsqu'on les place dans l'obscurité totale. Lors des périodes de repos, les insectes adoptent une posture caractéristique, tête et antennes repliées.
Les amphibiens et les poissons prennent également des postures typiques pendant leurs phases de repos. De plus, un nouveau critère a été récemment introduit qui consiste à priver un animal de ses périodes de repos pour voir si un rebond compensatoire - une augmentation de cet état - est observé lors de la récupération. Les insectes et les poissons présentent en effet un accroissement de cet état de repos profond après en avoir été sevrés, exactement comme les mammifères lorsqu'on les prive de sommeil. Mais si ces observations suggèrent qu'un état proche du sommeil peut exister chez ces animaux, nous ne disposons d'aucun indice de la présence d'un état voisin du sommeil paradoxal.
Les reptiles sont dans l'évolution voisins des mammifères. Leur cerveau présente de remarquables similitudes. C'est pourquoi beaucoup d'efforts ont été consacrés à l'étude électrophysiologique des états de vigilance chez les reptiles. Pourtant il manque une structure importante à leur cerveau pour permettre une électrogenèse proche de celle des espèces évoluées. Leur cortex cérébral n'est en effet présent que sous une forme très primitive (une couche fine), alors que l'essentiel de l'activité électrique que nous étudions pour définir le sommeil est généré par cette structure. Quoi qu'il en soit, les résultats sont souvent contradictoires; certains auteurs soutiennent l'existence d'un sommeil lent, voire même d'un sommeil paradoxal chez certains reptiles (les crocodiliens comme le caïman), tandis que d'autres nient toute modification de leur activité électrique cérébrale.
Cinq ans après la découverte du sommeil paradoxal chez le chat, l'équipe du neurobiologiste Michel Jouvet a pu démontrer l'existence du sommeil paradoxal, défini électrophysiologiquement, chez le pigeon et la poule. Deux autres particularités ont aussi été observées: chez les oiseaux, le sommeil paradoxal est particulièrement court, s'étalant de quelques secondes à une minute au maximum; les poussins ont une quantité plus conséquente de sommeil paradoxal que les adultes. L'existence indiscutable du sommeil paradoxal chez les oiseaux et sa quantité importante chez les oisillons approchent de près des mammifères, tandis que la brièveté des épisodes de sommeil paradoxal n'a pas encore trouvé d'explication satisfaisante. Plus récemment, un sommeil unilatéral a été observé chez certains oiseaux, avec des signes du sommeil sur un hémisphère et une fermeture de l'œil controlatéral. Cette dissymétrie pourrait expliquer les faibles durées de sommeil observées chez ces animaux. D'autre part, l'atonie musculaire du cou, l'une des caractéristiques propres aux mammifères, est rarement observée chez les oiseaux, à l'exception de l'oie.
L'existence du sommeil paradoxal chez l'ensemble des mammifères est une certitude, sauf pour deux espèces, le dauphin et l'échidné. Même si les mammifères ne représentent qu'une infime partie des espèces vivantes, ils sont les plus étudiés. Ainsi, le sommeil d'une centaine d'espèces de mammifères a déjà été polygraphiquement enregistré.
A quelques rares exceptions près, nous détectons l'existence du sommeil paradoxal uniquement chez les oiseaux et les mammifères, ce qui laisse à penser que le sommeil paradoxal est apparu très récemment au cours de l'évolution des espèces. Cette apparition coïncide avec celle de l'homéothermie, c'est-à-dire la capacité à maintenir une température centrale élevée et constante quelle que soit celle de l'environnement. Cependant, l'origine de l'homéothermie est incertaine et nous ne savons pas si les anciens reptiles thérapsidés, ancêtres des mammifères, ou les dinosaures étaient homéothermes ou pas. L'existence simultanée du sommeil paradoxal et de l'homéothermie ne nous apprend pas grand-chose quant à l'origine de chacune de ces fonctions car les mammifères et les oiseaux ont évolué séparément.
Ceci est vrai aussi pour leur absence simultanée chez les reptiles actuels, étant donné que les mammifères et les oiseaux, à l'opposé d'une croyance populaire, ne descendent pas de ces reptiles mais de formes beaucoup plus anciennes communes à tous. L'absence de liens phylogénétiques entre les grandes classes d'espèces actuelles nous empêche de déterminer à partir de quelle forme primitive le sommeil paradoxal et l'homéothermie ont évolué. En outre, nous savons que d'autres caractéristiques identiques entre les oiseaux et les mammifères, comme par exemple le cœurà quatre cavités, ont pu évoluer séparément sans qu'on puisse identifier les formes primitives communes. Il est également vrai qu'avec l'homéothermie disparaît la division cellulaire dans le système nerveux adulte tandis que cette fonction persiste chez les poïkilothermes. Encore une fois, ceci peut avoir évolué indépendamment du sommeil paradoxal et donc sans liens fonctionnels avec ce dernier.
Il existe une corrélation inverse entre le poids corporel et la quantité totale de sommeil. De la même façon, moins un animal dort, moins il présente de sommeil paradoxal. Toutefois, deux remarques s'imposent. Premièrement, cette corrélation ne se retrouve que chez les mammifères et les oiseaux, qui n'ont que très peu de sommeil paradoxal, et sont souvent de taille et de poids incomparables à ceux des mammifères. Deuxièmement, les plus gros oiseaux, comme le pingouin, ont plus de sommeil paradoxal que les plus petits. Enfin, ceci est vrai si l'on considère l'ensemble des mammifères car à l'intérieur de chaque ordre ou même d'une même espèce, la durée du sommeil paradoxal varie considérablement. Ainsi, le sommeil paradoxal diffère entre 1,5 et 5 heures chez les marsupiaux, 3,1 et 6 heures chez les édentés, 0,8 et 3,4 heures chez les rongeurs, 1,3 et 3,2 heures chez les carnivores et entre 0,7 et 2 heures chez les primates. Toutefois, l'absence de sommeil paradoxal chez les cétacés peut résulter de cette corrélation car il s'agit des plus gros mammifères.
Plus un animal est immature à la naissance plus il a du sommeil paradoxal; et ceci non seulement à la naissance mais également à l'âge adulte. C'est probablement la corrélation la plus forte trouvée pour expliquer la quantité de sommeil paradoxal. Or, encore une fois, cette corrélation n'est vérifiée que chez les mammifères, et les oiseaux, très immatures à la naissance, n'ont que peu de sommeil paradoxal. Toutefois, l'ontogenèse du sommeil paradoxal semble indiquer, aussi bien chez les oiseaux que chez les mammifères, que la quantité de sommeil paradoxal est environ cinq fois plus élevée à la naissance qu'à l'âge adulte.
Les animaux disposant d'un refuge sûr dorment et présentent davantage de sommeil paradoxal que ceux qui vivent dans des conditions précaires. Ainsi, l'homme et les animaux domestiques ont des quantités importantes de sommeil paradoxal, contrairement aux animaux victimes de prédateurs tels que le lapin. Ceci n'est pas vérifié chez les oiseaux, et la quantité de sommeil paradoxal ne semble pas être différente selon qu'il s'agisse de prédateurs, comme le faucon, ou de proies, comme l'étourneau, même si nous n'avons pas encore étudié un nombre suffisant d'espèces d'oiseaux. Le sommeil paradoxal du pingouin est important : plus d'une heure par jour, même s'il vit dans des conditions climatiques extrêmes (le froid est connu pour diminuer le sommeil paradoxal). Ceci peut s'expliquer par le fait que les pingouins, étudiés lors de la saison de reproduction, se tiennent toujours à une centaine de kilomètres des trous d'eau où vivent les prédateurs (phoques et baleines).
D'autres conditions écologiques peuvent être impliquées dans la quantité de sommeil paradoxal: la posture de sommeil, le froid ou encore la disponibilité de nourriture. La faible quantité de sommeil paradoxal chez les oiseaux ou même chez le babouin peut être en relation avec leurs postures nécessitant un certain niveau de tonus musculaire incompatible avec l'atonie du sommeil paradoxal. Toutefois, la quantité de sommeil paradoxal des oies, chez qui l'atonie totale du cou peut être observée lors de ces épisodes de sommeil, n'est pas différente de celles des autres oiseaux. L'exposition au froid diminue aussi notablement la quantité de sommeil paradoxal chez les mammifères, mais encore une fois les oiseaux semblent indifférents à la température ambiante (le froid n'a aucun effet sur la quantité de sommeil paradoxal chez le pingouin et le pigeon). En ce qui concerne la disponibilité de nourriture, les ruminants, qui ont besoin de quantités considérables d'herbe pour subvenir à leurs besoins alimentaires, dorment et présentent très peu de sommeil paradoxal lorsqu'ils sont en pâturage, mais dorment davantage et ont une quantité plus importante de sommeil paradoxal s'ils sont à l'étable avec un accès illimité à la nourriture.
Tous les êtres vivants y compris les unicellulaires ont des périodes journalières d'activité et de repos. Ceci reflète un rythme endogène réglé sur 24 heures (ou rythme circadien) dont nous commençons a comprendre les bases génétiques. Toutefois, nous manquons de critères physiologiques pour caractériser et distinguer les différentes phases de sommeils lent et paradoxal chez la plus grande majorité des animaux.
Dans l'état actuel de nos connaissances, l'étude phylogénétique du sommeil paradoxal ne semble pas apporter de réponses sûres quant à l'origine et l'évolution du sommeil paradoxal. Nous avons accumulé une grande quantité d'informations sur le sommeil des mammifères et avons cherché des corrélations pour tenter de comprendre les fonctions des sommeils lent et paradoxal mais ces informations sont partielles et n'expliquent pas le sommeil d'autres classes d'animaux. Des centaines d'autres espèces doivent êtres étudiées avant de se prononcer définitivement .
Le sommeil ne laissant pas de traces paléontologiques identifiées, nous ne saurons peut-être jamais à quel moment il s'est manifesté dans l'histoire de l'évolution des espèces. L'apparition du sommeil paradoxal chez les homéothermes et son association avec des rêves chez l'homme semble indiquer qu'il s'agit d'une fonction récente. Et pourtant, le dysfonctionnement de la thermorégulation pendant les phases de sommeil paradoxal et le fait que le pont, une des plus anciennes structures du cerveau, soit le siège de sa génération, semblent faire remonter son origine au temps des reptiles.
| L'émergence du sommeil paradoxal |
|
|
|
Le dysfonctionnement de la thermorégulation pendant les phases de sommeil paradoxal et le fait que le pont, une des plus anciennes structures du cerveau, soit le siège de sa génération, tendent à faire remonter son origine au temps des reptiles. |



| A quoi rêvent les bêtes ? |
 |
|
Toute personne ayant un chat ou chien vous dira qu'ils rêvent, car ces animaux présentent des comportements typiques de leur espèce, tout en étant endormis. Chez l'homme, le rêve s'élabore par la conjonction du sommeil paradoxal, de la mémoire et du langage. Nous savons que si l'on réveille un donneur pendant une phase de sommeil paradoxal et qu'on lui demande s'il rêvait, nous avons 80 % de probabilités de recueillir un rêve comportant des images et actions. C'est pourquoi les rêves ont été associés au sommeil paradoxal. Or, une activité mentale existe aussi pendant le sommeil lent, mais elle se différencie souvent du rêve par sa pauvreté en images. Si l'on admet que le sommeil paradoxal s'accompagne de rêves, tous les oiseaux et les mammifères qui ont du sommeil paradoxal, devraient également avoir du rêve. De la même façon, l'absence de sommeil paradoxal chez les autres animaux exclurait l'existence d'une activité onirique. Toutefois, les rêves utilisent des éléments mémorisés, aussi bien dans la mémoire à court terme que dans celle à long terme. Même si les structures cérébrales, nécessaires à la mémoire, existent chez une grande partie des animaux, nous connaissons très mal leur capacité de mémoire. Néanmoins, nous devons admettre que la grande majorité des animaux (à l'exception peut-être des insectes et des poissons) a probablement des traces mémorisées, nécessaires à la réalisation des rêves. Le dernier problème à résoudre reste le moyen de détection des rêves chez les animaux. Le seul outil est le langage, avec lequel nous communiquons le contenu de nos rêves. L'absence de langage chez tous les autres animaux nous interdit de connaître leurs rêves. Nous avons tout de même un moyen expérimental qui consiste à abolir l'atonie musculaire caractéristique du sommeil paradoxal, et à ainsi observer le comportement de l'animal pendant les phases de sommeil paradoxal. Dans ces conditions, un chat présente des comportements typiques de jeu ou de chasse d'un objet ou d'une souris imaginaire ainsi que des comportements d'agressivité et de défense. Tout laisse à penser qu'un chat, comme un homme, rêve de scénarii caractéristiques de son espèce pendant ses phases de sommeil paradoxal. M. T. |
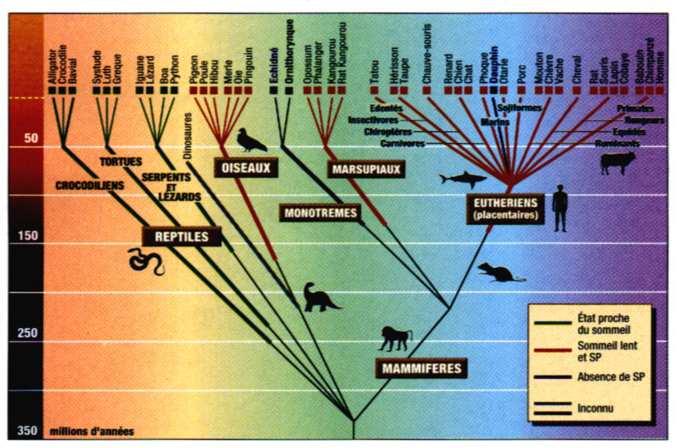
| Arbre phylogénétique... |
 |
|
...montrant les relations temporelles et les origines probables des reptiles, oiseaux et mammifères. Les lignes épaisses indiquent l'existence de fossiles permettant d'estimer les dates d'apparition des animaux, tandis que les lignes fines indiquent des relations phylogénétiques incertaines. Les reptiles (en vert) présentent des états de sommeil semblables au sommeil lent, mais il sont dépourvus de sommeil paradoxal. Celui-ci apparaît chez les oiseaux et les mammifères (en rouge), à deux exceptions près le dauphin et l'échidné. Par ailleurs, le sommeil de l'autre espèce de monotrème, l'ornithorynque (en noir), n'a pas encore été étudié. Enfin, le sommeil ne laissant pas de traces paléontologiques, nous ne saurons probablement jamais comment "dormaient" les dinosaures en gris), disparus il y a quelque 65 millions d'années. |
| Le sommeil primitif de l'échidné |
 |
|
L'échidné est l'un des deux représentants des monotrèmes encore vivants, l'autre étant l'ornithorynque. Ce sont les mammifères les plus anciens qui ont divergé des autres il y a environ 150 millions d'années. De ce fait, ils sont considérés comme des fossiles vivants. La première particularité de l'échidné est qu'il est ovipare, la seconde qu'il est un homéotherme imparfait: sa température cérébrale ne dépasse pas les 33°C et il peut hiberner. L'intérêt électrophysiologique qui lui est porté réside dans le fait qu'en 1972, Truett Allison a démontré l'existence du sommeil lent (S.L.) mais pas celle du sommeil paradoxal (S.P.). Or, s'agissant d'un mammifère fossile, on pense que l'étude de son sommeil doit nous renseigner sur les origines mêmes des différents types de sommeil. En fait, dès la première étude, un doute s'est installé sur l'existence du S.P. chez l'échidné car il présente des périodes de veille calme, sans tonus musculaire, accompagnées de mouvements oculaires semblables à ceux du S.P. Toutefois, la quasi-totalité des indices physiologiques, caractéristiques du S.P. des mammifères, est absente pendant ces périodes: le seuil d'éveil par stimulation est largement inférieur à celui du S.L., l'activité thêta hippocampique est absente, et le rythme cardiaque ne présente pas les irrégularités typiques du S.P. L'échidné est devenu le point de référence de la phylogenèse du S.P. et, au cours des récentes années, de beaucoup de théories sur la fonction du S.P. Son absence chez l'échidné ne peut pas être expliquée par ses particularités physiologiques comme sa température basse, car d'autres animaux avec des températures semblables ont du S.P., notamment le tatou qui est probablement l'animal en présentant le plus. L'échidné contredit aussi les corrélations qu'on trouve chez l'ensemble des mammifères pour expliquer la quantité de S.P., à savoir: plus un animal est petit et léger plus il a de S.P., or l'échidné est relativement petit; plus le refuge d'un animal est sûr plus il a du S.P., or l'échidné s'enfouit dans la terre et dort en totale sécurité; plus un animal est immature à la naissance plus il a du S.P., même à l'âge adulte, or le petit échidné est extrêmement immature à l'éclosion de l'œuf. Le mystère du sommeil de l'échidné commence peut-être à trouver un début de réponse, grâce aux travaux récents du professeur Siegel, aux États-Unis. Nous savons que l'activité électrique de certains neurones change de rythmicité et d'intensité, selon qu'un animal se trouve éveillé, en S.L. ou en S.P. Or, certains neurones, enregistrés directement dans le cerveau des échidnés, ont une activité électrique qui ne ressemble ni à celle du S.L. ni à celle du S.P. Pendant le sommeil de l'échidné, l'activité de ces neurones diminue comme pendant le S.L., mais sa variabilité augmente comme durant le S.P. Ainsi, il semble que l'échidné ne soit pas seulement dépourvu de S.P. mais également de S.L., comme celui défini chez les mammifères. Autre interprétation, le sommeil de l'échidné serait la forme primitive à partir de laquelle les deux sommeils ont évolué chez les mammifères. Si cette hypothèse s'avère exacte, nous aurons enfin trouvé un critère pour traquer les origines du sommeil et ses fonctions. Toutefois, il manque une dernière donnée à cette énigme. Nous savons que la quantité de S.P. diminue dramatiquement au cours de l'ontogenèse, les animaux adultes ayant environ cinq fois moins de S.P. que leurs nouveau-nés. Si des jeunes échidnés présentent très peu de S.P., ceci pourrait expliquer sa disparition chez les adultes. Des études sont encore nécessaires avant de se prononcer sur l'absence de S.P. chez cette espèce. M. T. |
| Le cerveau dédoublé du dauphin |
 |
|
Le sommeil des mammifères a évolué chez les animaux terrestres. Des adaptations sont mises en place chez les mammifères marins, la plus importante de celles-ci étant la nécessité de remonter à la surface pour respirer. Chez les dauphins, ainsi que chez certains autres cétacés et siréniens, le sommeil est unilatéral. Pendant qu'un hémisphère cérébral montre des signes typiques du sommeil lent, l'autre hémisphère reste éveillé. Comme la respiration est volontaire chez ces animaux, cette particularité leur permet d'assurer simultanément deux fonctions vitales: dormir et respirer; car dormir de tout son cerveau provoquerait l'arrêt respiratoire et la mort de l'animal. Le dauphin, qui ne s'arrête jamais de nager même en dormant, ne semble pas avoir de sommeil paradoxal (S.P.). Il s'agit d'une exception car les autres mammifères malins étudiés, comme les phoques et les otaries, ont un S.P. comparable à celui des mammifères terrestres. Toutefois, il est possible que les dauphins aient un S.P. unilatéral, impossible à détecter. Cette incapacité est liée au fait que les autres signes de S.P., comme l'absence de tonus musculaire et de mouvements oculaires rapides, ne sont pas présents chez ces mammifères. L'impossibilité à mettre en évidence le S.P. chez les dauphins peut aussi être due aux conditions expérimentales d'étude en milieu clos. Nous n'apportons ainsi encore aucune explication à cette énigme. M. T. |
| L'expérimentation onirologique |
 |
|
Les difficultés techniques d'étude de sommeil sont liées au fait que l'animal doit être préparé pour l'enregistrement polygraphique (EEG, EMG, EOG) pour être ensuite observé. Pour obtenir un rythme veille-sommeil proche de celui naturel d'un animal, l'idéal serait de faire l'expérimentation dans son milieu. Des techniques d'enregistrement télémétrique permettent d'équiper l'animal d'un émetteur, le libérant des câbles. Malheureusement, ces techniques ne sont pas généralisées et adaptées aux différentes espèces. Aussi, la quasi-totalité des études est réalisée au laboratoire. Même si l'enregistrement du sommeil nécessite une opération chirurgicale pour l'implantation d'électrodes, il s'agit d'une expérimentation animale parmi les moins traumatisantes. Toutes les précautions doivent être prises pour minimiser le stress et les manipulations invasives. Un animal ainsi préparé pour l'expérimentation vit normalement et en bonne santé pendant plus d'une année. L'étude du sommeil est effectuée par un spécialiste, souvent sous le contrôle des instances vétérinaires publiques et, sans exception, après l'obtention d'une autorisation d'un comité d'éthique qui vérifie la pertinence et la nécessité scientifique de l'expérimentation ainsi que les modalités strictes du respect des animaux. M.T. |
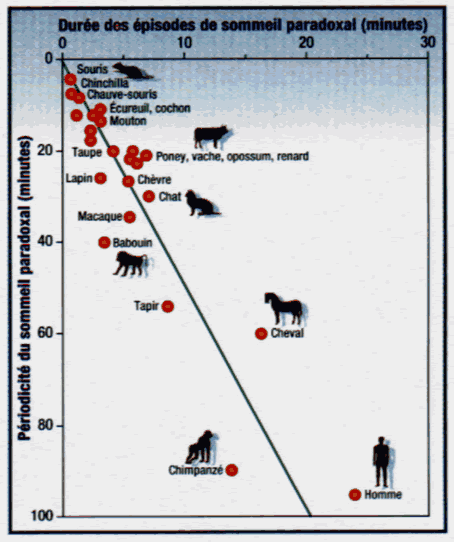
| La rythmicité du sommeil paradoxal |
 |
|
Le sommeil paradoxal n'est pas continu au cours du sommeil: il apparaît périodiquement et sa récurrence rythmique structure les cycles de sommeil. Sa périodicité est propre à chaque espèce animale: il se manifeste grossièrement toutes les 4 minutes de sommeil chez la souris, toutes les 12 minutes chez l'écureuil, toutes les 27 minutes chez le chat, toutes les 60 minutes chez le cheval, toutes les 90 minutes chez l'homme et toutes les 100 minutes chez l'éléphant. Ces cycles réguliers d'une heure et demie chez l'homme ont fait couler beaucoup d'encre. En 1963, Nathaniel Kleitman a imaginé que ce cycle pourrait refléter pendant la nuit un rythme physiologique d'alternance éveil/repos dans la journée, comme la partie émergée d'un iceberg. Il a été suivi sur cette voie par de nombreux scientifiques à la recherche d'un rythme originel de 90 minutes qui gouvernerait mobilité, performances, système végétatif et hormones. Ces chercheurs se sont cependant heurtés à de nombreuses contradictions expérimentales, car ce rythme varie d'un individu à l'autre, d'une nuit à l'autre et même à l'intérieur d'une même nuit, avec des écarts atteignant 50 minutes. Comme d'autres grandeurs physiologiques telles que la durée des cycles cardiaques, respiratoires ou de gestation, la périodicité du S.P. est inversement proportionnelle, à l'exception des ruminants, au métabolisme de base de ces animaux. Ainsi la souris, qui a un métabolisme de base 25 fois plus grand que l'éléphant (c'est-à-dire qu'elle consomme vingt cinq fois plus d'oxygène par gramme de poids et par heure), présente 25 fois plus souvent du S.P. que l'éléphant... Si la souris rêve plus fréquemment, elle rêve moins longtemps que l'éléphant. C'est ainsi que si nous comparons la durée d'un épisode de S.P. avec sa périodicité chez les 26 espèces animales pour lesquelles des données fiables sont disponibles, nous trouvons un rapport, dit "rapport cyclique", extrêmement fixe et proche de 4. Cela signifie que chaque cycle de sommeil comporte trois quarts de sommeil lent suivis d'un quart de S.P., et ce dans toutes les espèces animales. A cette périodicité qui semble déterminée par le métabolisme de base de notre espèce, s'ajoutent des caractéristiques énergétiques cérébrales opposées entre sommeil lent et S.P. Ainsi, pendant le sommeil lent, les neurones corticaux qui fonctionnent de façon synchrone en quelque sorte au ralenti, réduisent d'un tiers leur consommation de glucose et d'oxygène. A l'inverse, en S.P., le cerveau se gorge de sang et les neurones hyperactifs consomment des quantités de glucose et d'oxygène telles qu'elles peuvent dépasser les valeurs de l'éveil. Un sommeil lent économe en énergie pour notre cerveau alterne ainsi régulièrement avec un S.P. luxueux et consommateur d'énergie. Les situations de carence énergétique cérébrale, telles que l'hypoglycémie, l'hypoxie, l'épilepsie et la fièvre s'accompagnent d'une réduction, voire d'une suppression du S.P. Il serait ainsi possible que des conditions énergétiques cérébrales favorables conditionnent l'apparition cyclique du S.P. Pour mieux comprendre cette relation entre métabolisme énergétique cérébral et périodicité du S.P., nous avons pu modifier dans des conditions expérimentales très particulières la périodicité du S.P. chez le chat en donnant à respirer à ces animaux de l'oxygène pur. Le S.P. est alors apparu toutes les 30 minutes, soit deux fois plus souvent qu'avec de l'air normal. C'est un peu comme si l'on demandait subitement à une horloge de tourner deux fois plus vite, en lui fournissant en quelque sorte l'énergie pour le faire. C'est la première fois que l'on peut modifier de façon claire et reproductible le cycle du S.P., alors que ni les médicaments, ni l'exposition à des lumières discontinues ou continues, ni l'isolement temporel n'affectent cette périodicité. Ainsi, le métabolisme énergétique de l'animal, qu'il soit cérébral ou corporel, semble influencer fortement la périodicité de son S.P. Isabelle Arnulf, |
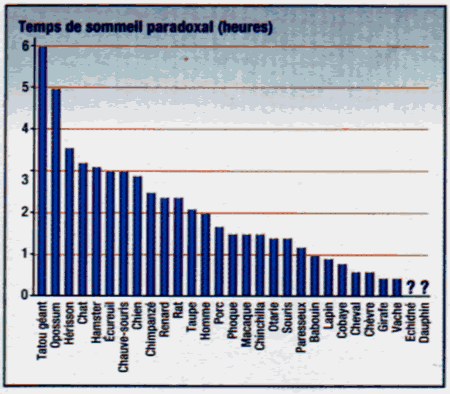
| Gros et petits "rêveurs" |
 |
|
Malgré l'importance que certains scientifiques donnent au sommeil paradoxal pour le développement des fonctions supérieures du cerveau, les deux mammifères les plus primitifs (le tatou géant et l'opossum) présentent des quantités impressionnantes de sommeil paradoxal (S.P.), tandis que l'homme n'en a qu'une quantité très moyenne. En général, plus un animal dort, plus il présente de S.P.. Ainsi, le tatou géant et l'opossum sont de très gros dormeurs (18 heures par jour), tandis que les ruminants, comme la vache, sont de très petits dormeurs (environ 4 heures par jour). Il y a toutefois des exceptions à cette règle car le paresseux par exemple, qui peut dormir jusqu'à 17 heures par jour n'a que 72 minutes de S.P. M. T. |