| Documents neurophysiologiques relatifs aux mécanismes de l'attention chez l'homme |
| Par M. Jouvet *, B. Schott, J. Courjon Et G. Allegre Revue Neurologique Tome 100 N°5 1959 (pp. 437-450) |
* Chargé de recherches an C.N.R.S.
(1) Travail de la Clinique Neuropsychiatrique de la Faculté de
Médecine de Lyon (Pr Dechaume)
Ce travail a bénéficié d'une aide de l'E.O.A.R.D.C. - U.S.A.F. - Contrat AF 61 (514) 1206
Parmi les nombreuses modalités de réaction de l'organisme aux différents messages du milieu extérieur, l'attention constitue un phénomène privilégié ; elle traduit en effet la réponse organisée et totale du système nerveux à un signal important. Plus que la réactivité à un signal, l'attention objective la perceptivité de celui-ci. On conçoit donc l'intérêt qu'il peut y avoir à étudier les corrélations neurophysiologiques de ce processus afin de mieux comprendre les mécanismes de la prise de conscience. L'expérience subjective de chaque jour permet de reconnaître les deux aspects fondamentaux du processus d'attention que la psychologie a bien mis en évidence, c'est dire, la sélectivité de l'attention pour une seule catégorie de signaux et l'effet inhibiteur qui s'exerce sur les autres modalités sensorielles. Pour le neurophysiologiste deux problèmes sont ainsi à résoudre :
- le premier concerne l'aspect positif de l'attention : quelle va être au sein du système nerveux la répartition et l'allure des signaux qui conditionnent l'attention ?
- le second s'applique à l'aspect négatif de ce phénomène, la distraction en quelque sorte - c'est le devenir des autres messages du monde extérieur devant ce processus. Adrian (1) remarque à ce sujet que les influx des organes sensoriels doivent être "intégrés" de façon différente lorsque nous y sommes attentifs et lorsque nous ne le sommes pas. Pour cet auteur deux mécanismes sont possibles :
Dans le premier, tous les signaux parviendraient au cortex, quel que soit l'objet de l'attention. Le cortex aurait ainsi à sa disposition à chaque instant un modèle complet de l'univers environnant transmis par les récepteurs sensoriels et il reviendrait à un mécanisme de contrôle de rendre une aire corticale privilégiée réceptive au signal auquel nous sommes attentifs.
Selon un autre mécanisme, l'appareil de contrôle interviendrait à un stade plus précoce, et seuls les messages privilégiés parviendraient au cortex.
L'expérimentation animale s'est efforcée récemment (13-14) d'objectiver les mécanismes essentiels de l'attention, mais il s'agit d'une expérimentation difficile. Il est parfois impossible en effet, en expérience aiguë sur un animal curarisé et non anesthésié ou en expérience chronique, de distinguer par l'analyse de l'activité électrique corticale ou du comportement un état d'attention d'un simple éveil cortical non spécifique. C'est pourquoi l'étude de la transmission des messages corticipètes chez l'homme prend une importance particulière - la connaissance de l'expérience subjective du sujet humain est infiniment précieuse pour l'expérimentateur qui peut ainsi contrôler la réalité du processus d'attention qu'il induit.
De son côté la clinique neurologique offre l'occasion quotidienne d'observer des troubles de l'attention, que ceux-ci traduisent une dissolution globale de la conscience ou au contraire qu'ils objectivent une atteinte à l'emporte-pièce dans le champ de la conscience. Ces cas privilégiés, véritable "dissolution de conscience en secteur " comme les nomme Purdon-Martin constituent de "véritables expériences faites sur le cerveau par la maladie" selon le mot de H. Jackson. Et l'on conçoit l'intérêt qu'il peut y avoir à explorer de la manière la plus objective possible cette pathologie de l'attention.
C'est à l'E.E.G. recueilli sur le scalp que l'on s'est d'abord adressé, mais les renseignements ont été minimes. L'E.E.G. en effet n'est qu'un lointain reflet de l'activité électrique cérébrale et doit compter avec les perturbations liées aux lésions responsables du syndrome pathologique. D'autre part même si avec les méthodes d'intégration électroniques actuelles (3-4-5) on parvient à mettre en évidence les réponses corticales aux signaux afférents, rien ne permet d'affirmer, devant une variation d'amplitude, si celle-ci s'est produite à une étape précorticale ou au niveau du cortex.
Le but de nos investigations poursuivies depuis 2 ans (6) a été de vérifier s'il était possible d'objectiver chez l'homme un mécanisme de contrôle des signaux afférents au cours de leur étape précorticale lors de l'attention. Cette condition ayant été vérifiée, nous avons ensuite étudié des sujets atteints de troubles divers de la conscience. Nous exposerons d'abord les résultats portant sur les afférences visuelles et somesthésiques obtenus sur des groupes de sujets indemnes de troubles de l'attention, ensuite sur un groupe de sujets pathologiques.
I. - LES RÉPONSES VISUELLES SOUS-CORTICALES CHEZ L'HOMME
Technique.
Les résultats portent sur 8 sujets. Ces 8 sujets devaient subir une ventriculographie gazeuse (5 fois pour localiser une tumeur hémisphérique limitée, n'ayant pas donné d'image convaincante à l'artériographie, 3 fois pour un syndrome d'hypertension intracrânienne qui se révéla être une encéphalite pseudo-tumorale). Chez ces sujets, conscients, sous anesthésie locale, au niveau de l'orifice de trépanation occipItal.e fut fixé un appareil stéréotaxique léger (7) et une électrode bipolaire concentrique fut introduite dans le lobe occipItal. droit au niveau des radiations optiques. L'enregistrement sous-cortical et celui du scalp furent effectués sur un appareil E.E.G. Alvar.
La stimulation lumineuse intermittente est produite par un appareil électronique (Soneclat-Alvar) placé à 50 cm de la tête du sujet et délivrant des éclairs lumineux de forte intensité qui sont enregistrés sur une cellule photo-électrique fixée contre les deux yeux du sujet.
Ces enregistrements ont nécessité deux ordres de contrôle :
a) Le facteur périphérique oculaire.
Les pupilles du sujet sont dilatées de façon systématique avant l'enregistrement par de l'Atropine locale, et les paupières demeurent volontairement closes durant l'enregistrement. Un observateur placé à côté du sujet vérifie constamment l'absence de tous mouvements des paupières. Dans deux cas enfin a été réalisé l'enregistrement des mouvements des yeux au moyen d'électrodes placées au-dessus et au-dessous de chaque oeil selon la disposition décrite par A. Rémond (19). Il a été vérifié que les mouvements des globes oculaires n'entraînent pas de variations des réponses visuelles sous-corticales à des éclairs de forte intensité lumineuse. De plus, il fut aussi vérifié que les variations des réponses sous-corticales ne s'accompagnent qu'inconstamment de mouvements oculaires minimes.
b) Position de l'électrode d'enregistrement sous-corlical.
Cette position est vérifiée parla prise de radiographies de face et de profil après injection des ventricules cérébraux par de l'air au cours de la ventriculographie préopératoire qui fait suite à l'enregistrement.
Résultats.
Les réponses visuelles sous-corticales " témoins " sont obtenues clans un silence total, sans que l'on ait demandé au sujet de faire aucun effort d'attention et après qu'une certaine accoutumance à la situation expérimentale a été réalisée.
Ces réponses se traduisent par des potentiels évoqués complexes comportant une onde rapide diphasique initialement positive suivie d'une oncle lente diphasique initialement négative. Ces réponses sont analogues à celles décrites chez l'homme par d'autres auteurs (9-22) (fig. 1).
La fréquence maximum avec laquelle les réponses peuvent être entraînées par la stimulation lumineuse intermittente varie de 20 à 30 éclairs /seconde, selon les sujets. Ces réponses présentent enfin un certain degré de fluctuation d'amplitude qu'il est difficile de relater à un état subjectif du sujet.
Le moindre bruit dans la salle d'opération, une seule parole prononcée entraînent immédiatement une diminution nette de l'amplitude des réponses.
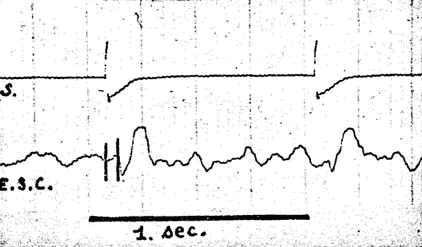
Fig. 1. - Réponse visuelle des radiations optiques du lobe occipItal. ; la latence est d'environ 50 millisecondes. Dans les figures 1, 3, 6, 9, 10, E.S.C. et E.S.G. représentent l'enregistrement sous-cortical ; R.O.B. et R.O.M. (fig. 7-8), même dérivation en bipolaire ou monopolaire. S, cellule photo-électrique objectivant le flash lumineux. Calibrage : 1 s 50 microvolts.
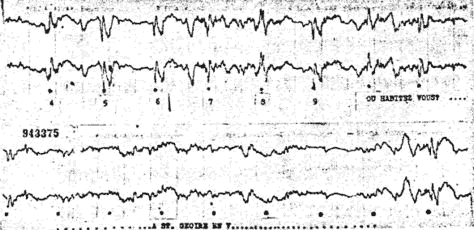
Fig. 2. - Augmentation d'amplitude des réponses occipItal.es lorsque le sujet compte les flashes (chiffres). Diminution des réponses lors d'attention auditive et d'évocation mnémonique.
Les réponses visuelles lors d'état d'attention visuelle.
Cette condition est réalisée en demandant au sujet de compter les éclairs lumineux (présentés à la fréquence d'un éclair /seconde environ). L'attention visuelle s'accompagne des corrélations E.E.G. suivantes:
- Activité rapide et de bas-voltage sur le scalp, surtout nette en dérivation occipItal.e - l'activité rapide s'observe en général au début de l'état d'attention. Elle laisse place rapidement au retour du rythme alpha.
- Augmentation d'amplitude des potentiels évoqués sous-corticaux portant sur les deux composantes rapides et lentes des réponses. Cette augmentation d'amplitude est constante pendant toute la durée de l'état d'attention (fig. 2).
Les réponses visuelles lors d'état d'attention portant sur d'autres modalités sensorielles (distraction).
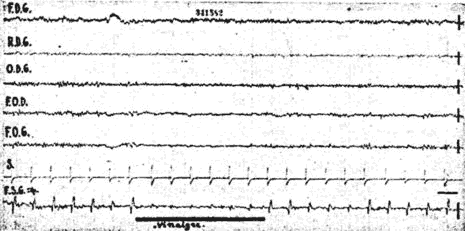
Fig. 3. - Blocage des réponses visuelles sous-corticales lors d'attention olfactive (reconnaissance d'une odeur de vinaigre). Il existe en même temps une discrète réaction d'arrêt sur le scalp.
Ce phénomène s'accompagne d'une réduction d'amplitude des réponses visuelles, pouvant parfois aller jusqu'à leur disparition presque totale. Ce "blocage" des potentiels évoqués sous-corticaux visuels a été observé de façon très nette lors de stimulations nociceptives (piqûre de la cuisse, déclenchement spontané d'une crise de névralgie du trijumeau chez une malade).
Les états d'attention olfactive (reconnaissance d'une odeur) ont entraîné également des blocages importants (fig. 3). Lors des états d'attention auditive les questions de l'expérimentateur entraînent une diminution d'amplitude des réponses plus constantes et plus intenses que des états d'attention déclenchés par des stimuli plus simples. (écouter le bruit d'une montre par exemple).
C'est l'attention somesthésique (reconnaissance d'un objet placé dans une main) qui a entraîné les blocages les moins importants.
Il en est de même des processus " mentaux ", certains sujets ont présenté des diminutions importantes d'amplitude des réponses visuelles lors de problèmes simples (date de naissance, nom du Président de la République, calcul mental) alors que chez d'autres les mêmes questions n'entraînaient aucune variation d'amplitude ; en général le calcul mental simple (table de multiplication) n'entraîne aucun "blocage". Par contre, les additions de nombre de 3 chiffres s'accompagnent de blocage très bref au fur et à mesure que les sujets "posent les retenues".
Il n'a pas toujours été possible de trouver une corrélation étroite entre les blocages sous-corticaux et une activité rapide au niveau du scalp, soit généralisée, soit localisée.
II. - LES RÉPONSES THALAMIQUES SOMESTHÉSIQUES
La première observation concerne un sujet masculin, âgé de 72 ans, porteur d'un carcinome prostatique avec envahissement du plexus sacré entraînant des douleurs intolérables au niveau du membre inférieur gauche. L'intensité des douleurs, leur résistance au traitement médical font poser l'indication d'une destruction localisée thalamique. Sous anesthésie locale, une électrode multipolaire servant à l'enregistrement, la stimulation et coagulation est introduite de façon stéréotaxique au niveau du noyau ventro-postéro-latéral du thalamus droit (V. P. L.) n, après repérage ventriculographique selon la technique et avec l'appareil de Talairach (20). Les stimuli somesthésiques sont produits par un aide qui. percute rythmiquement les téguments au moyen d'une pince, avec une force constante, suffisante pour évoquer une sensation de percussion non douloureuse. L'enregistrement électrique est effectué par un appareil E.E.G. La stimulation est obtenue avec un stimulateur Grass.
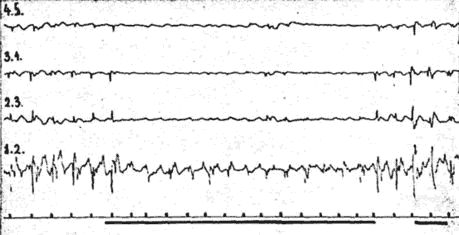
Fig. 4. - Réponses somesthésiques thalamiques (noyau ventro-postéro-latéral droit). L'électrode la plus inférieure, recueille les potentiels évoqués par simulation mécanique de l'hémiface gauche. Blocage des réponses lors de calcul mental (132 x 6) (marqué par le trait noie).
Une fois le V.P.L. repéré, une coagulation a été effectuée à son niveau. Les suites opératoires ont été bonnes. Le malade n'a pas présenté d'anesthésie véritable mais une agnosie douloureuse du type de celle que l'on observe après leucotomie.
La deuxième exploration a été effectuée sur un sujet masculin de 52 ans, souffrant, d'algies faciales récidivantes après neurectomie rétrogassérienne itérative. Dans ce cas notre appareil stéréotaxique fut employé ; les mêmes conditions de stimulation et d'enregistrement furent mises en jeu. Après thalamotomie, ce malade opéré il y a deux mois, se déclare très soulagé et a repris plusieurs kilogrammes.
Résultats.
Les réponses somesthésiques thalamiques :
Celles que nous décrivons ont été obtenues par la percussion d'une zone limitée de l'hémiface gauche (partie inférieure de la joue, menton). La stimulation au niveau de l'électrode d'enregistrement (1 /seconde, 10 millisecondes, 4 volts) a provoqué des sensations légères au niveau de cette même région en même temps qu'elle entraînait de légères clonies des muscles de cette zone. L'allure de ces réponses est complexe. La première phase négative étant souvent entrecoupée de déflexion positive. En raison des conditions d'enregistrement, nous n'avons pu étudier leur latence ni leur seuil. Seules ont été étudiées leurs variations systématiques en fonction de l'état d'attention du sujet conscient.
Lorsque l'attention du sujet se porte sur le stimulus somesthésique en lui demandant de "bien faire attention" et de compter les coups, l'amplitude des réponses augmente de façon très nette. Par contre, lors d'état de distraction (calcul mental - calculer mentalement 132 x 6) l'amplitude des réponses a diminué de façon très significative (fig. 4).
III. - EXPLORATION PORTANT SUR DES SUJETS ATTEINTS DE TROUBLES DE LA CONSCIENCE
Résumé des observations :
Obs. 1.
- Andrée P.... 30 ans, pas d'antécédent pathologique. Début de l'histoire clinique en novembre 1956, par des céphalées, des douleurs diffuses, de la fièvre, bientôt suivies d'un état de mai comitial, et d'un coma. Trachéotomie. Réaction cellulaire du L. C.-R. Depuis cette date, c'est-à-dire pendant 900 jours environ cette femme est restée hospItal.isée dans la clinique dans un état particulier de conscience caractérisé par :
- Une absence totale de perceptivité visuelle ou auditive.
- L'absence de clignement à la menace, de fixation des objets mouvants.
- Une réactivité à la douleur caractérisée par une mimique très riche, lors des stimulations nociceptives, une réaction d'orientation conservée lors de l'état de veille tourne la tête du côté d'une stimulation auditive forte.
- Absence totale de troubles végétatifs - état général excellent, aucune escarre.
Au point de vue moteur :
Membres inférieurs en hyperextension, membres supérieurs
en hyperflexion. Réflexe de Magnus et de Kleijn (cf. fig.
5).
Des investigations sous-corticales furent pratiquées sur cette
malade, respectivement au 200e et au 500e jour de son état pathologique.
La ventriculographie concomitante mit en évidence une importante
et progressive dilatation ventriculaire, sans hypertension liquidienne.
Résultats.
- Les réponses visuelles sous-corticales étaient identiques en tout point à celles des sujets normaux. La seule différence notable étant la présence de pointes rapides accompagnant les mouvements oculaires comparables aux ondes lambda.
Aucune variation d'amplitude ne put être constatée, malgré les stimulations nociceptives importantes entraînant des mimiques douloureuses (fig. 6).
Il semble donc que dans ce cas la fonction de transmission rétino-corticale soit intacte, mais que la lésion probable du tronc cérébral rende impossible tout blocage.
Obs. 2.
- Odette P.... 39 ans. Pas d'antécédents pathologiques notables. Début Subit de l'affection en a), ri! 1957 par un coma qui persiste trois jours, laissant place à un tableau d'atteinte du tronc cérébral, caractérisé par des troubles de la conscience évoquant le mutisme akinétique - absence totale de mouvements spontanés - mutisme, mais possibilité de réponse après des sollicitations longuement répétées.
Un syndrome neurologique associant une paralysie de l'élévation des yeux vers le haut, des spasmes de convergence, un nystagmus disjonctif, une aréflexie pupillaire à la lumière. Ces troubles persistent pendant quatre mois puis régressent ensuite lentement, laissant place à l'évolution d'une paraparégie spasmodique.
L'exploration sous-corticale fut pratiquée un mois après le début des troubles la ventriculographie montra une dilatation importante des ventricules.
Résultats.
Chez cette malade, les questions de l'expérimentateur (essai d'interprétation d'une odeur) n'entraînèrent pas de diminution des réponses. Par contre, des stimulations nociceptives entraînèrent un blocage fugace mais indéniable (fig. 7-8). Il semble donc que dans ce cas de mutisme akinétique, les troubles de la conscience portent surtout sur l'étape efférente des réponses puisque l'on peut objectiver l'un des aspects majeurs de l'attention c'est-à-dire le blocage.
Fig. 5. - Malade de l'observation 1. Réflexe de Magnus et Kleijn.
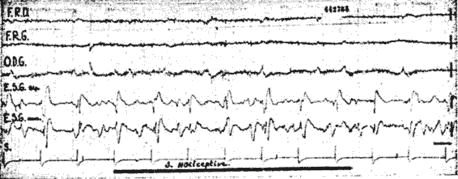
Fig. 6. - Absence de blocage des réponses visuelles sous-corticales lors d'une stimulation nociceptive. Malade de l'observation 1.
Obs. 3.
- Monique P..., 22 ans. Présente depuis l'âge de 16 ans de nombreuses manifestations de nature pithiatique : grande crise avec manifestations théâtrales, rétention d'urine durable, crises douloureuses abdominales ayant cédé chaque fois à la psychothérapie.
Rentre dans le service en juin 1957 en se plaignant de céphalées intolérables. L'examen révèle une anesthésie globale de tout le corps. La malade ne réagissant à aucune stimulation somesthésique, même la plus douloureuse, et déclarant ne rien sentir.
L'intensité des céphalées, l'aspect dysmorphique du crâne font poser l'indication d'une ventriculographie qui objective des ventricules normaux et symétriques. L'exploration sous-corticale a lieu à cette occasion. Cette malade quitte le service en juillet 1957 guérie de son dernier épisode, avec une sensibilité normale.
Résultats.
- Chez cette malade, aucune stimulation nociceptive : piqûre de la cuisse, torsion du gros orteil, n'entraîna de réaction clinique ou mimique. Elle déclarait ne ressentir aucune douleur. Parallèlement il ne fut observé observé aucune diminution de l'amplitude des réponses sous-corticales visuelles (voir fig. 9).
Par contre, à un certain moment, l'évocation d'un récent voyage qu'elle fit à Lourdes, et qui l'avait particulièrement impressionnée, entraîne un blocage fugace mais indéniable (voir fig. 10).
Cette malade fut ensuite guérie par psychothérapie aidée par des narcoses à l'Eunoctal.
Il semble donc, chez cette grande hystérique, que son anesthésie totale s'accompagnât d'un trouble fonctionnel portant sur l'attention à la douleur responsable du défaut de blocage observé.
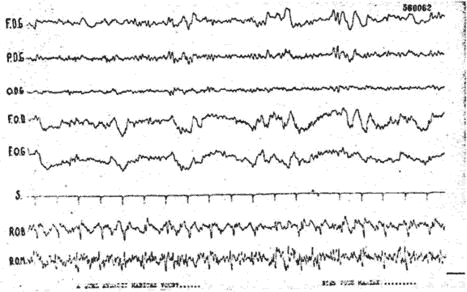
Fig. 7. - Malade de l'observation 2. Absence de blocage lors de l'attention auditive: question de l'expérimentateur. Remarquer l'aspect pathologique du tracé E.E.G.
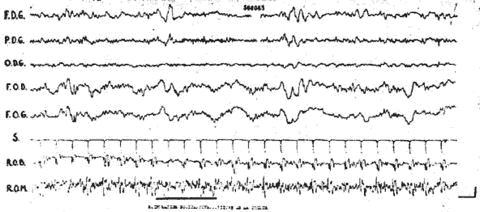
Fig. 8. - Malade de l'observation 2. Blocage discret, mais certain, au cours d'une stimulation nociceptive (piqûre de la cuisse).
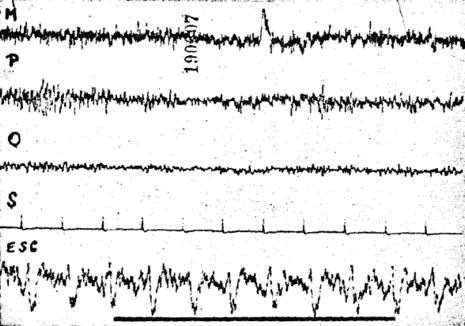
Fig. 9. - Malade de l'observation 3. Absence de blocage par stimulation nociceptive chez une hystérique atteinte d'anesthésie globale.
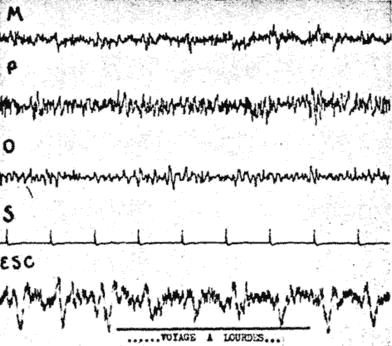
Fig. 10. - Malade de l'observation 3. Blocage discret lors de l'évocation d'un voyage récent à Lourdes.
DISCUSSION
L'attention au signal visuel ou somesthésique s'accompagne donc d'une augmentation relative d'amplitude du message corticipète. Cette facilitation des réponses permet ainsi de penser que le signal " attractif " parvient au cortex avec sa plus grande amplitude. Cette donnée confirme les résultats obtenus chez l'animal chronique lors d'état d'attention provoquée (15-16).
1. Cette augmentation d'amplitude du message précortical doit également être rapprochée des effets de facilitation des réponses afférentes sensorielles observées par certains auteurs par stimulation de la formation réticulaire du tronc cérébral (8).
2. La dépression des réponses corticipètes visuelles ou somesthésiques, lors d'états d'attention déclenchés par d'autres modalités sensorielles pose quelques problèmes.
Le blocage sous-cortical observé lors de l'attention oblige à faire une distinction entre les mécanismes neuroniques mis en jeu lors de la réaction d'arrêt et ceux mis en jeu lors de l'attention. Une dépression des réponses évoquées uniquement à l'étage cortical, sans blocage sous-jacent a en effet été constatée en expérimentation animale lors d'activation corticale provoquée par stimulation de la formation réticulaire mésencéphalique (2) par des stimulations sensorielles (15). On ne peut donc toujours assimiler la réaction d'arrêt de Berger au processus actif de l'attention. La réaction de " blocage activation" n'est ainsi ni la condition nécessaire (puisque dans nos cas nous avons observé des blocages sous-corticaux alors que le rythme alpha persistait), ni la condition suffisante du processus d'attention. Le blocage des messages sensoriels corticipètes confirme également les résultats déjà obtenus chez l'animal (13) mais il nous est impossible de préciser à l'heure actuelle à quel niveau s'effectue chez l'homme l'inhibition des messages afférents visuels (corps. genouillé externe, ou rétine) ou somesthésique (thalamus, moelle).
Ces données font donc admettre l'hypothèse soulevée par Adrian d'un mécanisme de contrôle intervenant dès l'étape afférente sous-corticale lors de l'attention. Certaines parties de la formation réticulée mésencéphalique jouent certainement un rôle important au sein de ce mécanisme de contrôle.
On sait, en effet, que la stimulation de la formation réticulée est capable de contrôler la transmission de messages sensoriels afférents musculaires (11), somesthésiques (12), visuels (10-14) ou auditifs (18). L'absence de blocage observée dans notre observation 1, où une lésion du tegmentum mésencéphalique est très probable, confirme ces données.
Cette mise en jeu des formations centrales responsables du blocage pourrait se faire à partir du cortex. La stimulation d'aires corticales en relation étroite avec la formation réticulée peut en effet "bloquer" certaines afférences au niveau des premiers relais spécifiques (17).
D'une façon générale, les résultats obtenus démontrent le rôle important joué par le cortex lors du processus d'attention puisque les messages importants lui parviennent avec leur amplitude maximale. Ils évoquent également la mise en jeu des formations multisynaptiques du trône cérébral clans le processus concomitant de blocage des autres modalités sensorielles (fig. 11).
Ce couplage fonctionnel de l'écorce et des formations réticulaires centrales paraît ainsi être une (les conditions nécessaires de l'attention, et par là de la prise de conscience d'un signal.
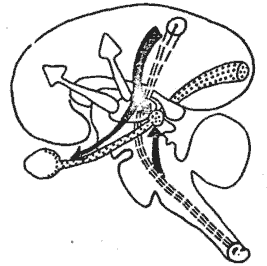
Fig. 11. - Schéma de mécanismes hypothétiques entrant en jeu au cours de l'attention. Le stimulus attractif somesthésique met enjeu des mécanismes inhibiteurs (flèches noires) à point de départ sous-cortical ou cortical, agissant sur les afférences visuelles (pointillé) soit au niveau rétinien, soit au relais géniculé. Il met en jeu également un mécanisme dynamogénique qui active l'écorce cérébrale (flèche blanche).
CONCLUSIONS
1. Sur le plan neurologique, nous lie voulons pas reprendre sur un seul cas la discussion des mécanismes responsables de l'hystérie. On sait d'ailleurs que les avis des électroencéphalographistes sont partagés à ce sujet. Il semble bien, cependant, qu'il faille admettre la réalité d'un trouble fonctionnel portant sur la perceptivité des signaux douloureux dans le cas de l'anesthésie hystérique. Mais à l'heure actuelle, il est bien difficile d'explorer un tel trouble fonctionnel.
2. A la lumière de nos résultats, les mécanismes de l'attention se laissent difficilement localiser, mais on peut se demander si l'étude systématique des signaux afférents corticipètes ne mettrait pas en évidence des troubles dynamiques dans le cas de troubles de l'attention localisés (syndrome d'Anton-Babinski) ou au contraire dans le cas d'exagération pathologique de l'attention décrit par Tournay (21).
3. La réalité d'un contrôle des afférences somesthésiques au niveau thalamique pose le problème de la douleur. Si nos observations concernent des signaux tactiles, on peut logiquement supposer que des signaux douloureux auraient subi un contrôle identique lors du relais thalamique. Cette possibilité évoque certains procédés d'analgésie. L'accouchement sans douleur, par exemple, où l'attention de la parturiente continuellement sollicitée par le contrôle de la respiration, par les encouragements des accoucheurs, viendrait bloquer les influx douloureux d'origine pelvienne.
4. Enfin nos résultats montrent que l'attention constitue une des conditions essentielles de la perceptivité d'un signal, chez la malade de l'obs. 1, en effet, aucune variation dynamique ne peut être objectivée sur des réponses apparemment normales. Cette malade n'est capable d'aucune perceptivité visuelle, même élémentaire car elle ne cligne pas à la menace. Par contre, sa réactivité à la douleur semble mettre en jeu des mécanismes dépendant d'un niveau plus bas, puisqu'ils sont incapables d'agir sur les réponses visuelles. Cette dissociation oblige à admettre deux niveaux fondamentalement différents : celui de la perception consciente d'un signal au cours duquel intervient le processus d'attention, celui de la réactivité mimique ou motrice, purement moteur, sans que soit mis en jeu le contrôle des afférences.
Cette dissociation est également manifeste dans le cas du mutisme akinétique (obs. 2), dans ce cas, la perceptivité est conservée, et c'est l'étape motrice efférente du comportement qui est perturbée. Il est classique d'ailleurs de signaler que ces sujets sont capables à certains moments de réponses fugitives montrant qu'ils sont parfaitement conscients.
Quelque fragmentaires qu'ils soient, et il ne peut en être autrement en Neurophysiologie clinique, ces documents permettent - nous l'espérons - de combler un peu le fossé qui sépare encore actuellement l'étude expérimentale de l'activité nerveuse supérieure chez l'animal des mécanismes de la prise de conscience chez l'homme.