| Documents neurophysiologiques relatifs aux mécanismes de l'attention chez l'homme |
| Par M. Jouvet *, B. Schott, J. Courjon Et G. Allegre Revue Neurologique Tome 100 N°5 1959 (pp. 437-450) |
| TABLE DES MATIERES |
|
III. - Exploration portant sur des sujets atteints de troubles de la conscience |
| IMPRESSION |
| Version imprimable (Tout l'article dans une seule page) |
DISCUSSION
L'attention au signal visuel ou somesthésique s'accompagne donc d'une augmentation relative d'amplitude du message corticipète. Cette facilitation des réponses permet ainsi de penser que le signal " attractif " parvient au cortex avec sa plus grande amplitude. Cette donnée confirme les résultats obtenus chez l'animal chronique lors d'état d'attention provoquée (15-16).
1. Cette augmentation d'amplitude du message précortical doit également être rapprochée des effets de facilitation des réponses afférentes sensorielles observées par certains auteurs par stimulation de la formation réticulaire du tronc cérébral (8).
2. La dépression des réponses corticipètes visuelles ou somesthésiques, lors d'états d'attention déclenchés par d'autres modalités sensorielles pose quelques problèmes.
Le blocage sous-cortical observé lors de l'attention oblige à faire une distinction entre les mécanismes neuroniques mis en jeu lors de la réaction d'arrêt et ceux mis en jeu lors de l'attention. Une dépression des réponses évoquées uniquement à l'étage cortical, sans blocage sous-jacent a en effet été constatée en expérimentation animale lors d'activation corticale provoquée par stimulation de la formation réticulaire mésencéphalique (2) par des stimulations sensorielles (15). On ne peut donc toujours assimiler la réaction d'arrêt de Berger au processus actif de l'attention. La réaction de " blocage activation" n'est ainsi ni la condition nécessaire (puisque dans nos cas nous avons observé des blocages sous-corticaux alors que le rythme alpha persistait), ni la condition suffisante du processus d'attention. Le blocage des messages sensoriels corticipètes confirme également les résultats déjà obtenus chez l'animal (13) mais il nous est impossible de préciser à l'heure actuelle à quel niveau s'effectue chez l'homme l'inhibition des messages afférents visuels (corps. genouillé externe, ou rétine) ou somesthésique (thalamus, moelle).
Ces données font donc admettre l'hypothèse soulevée par Adrian d'un mécanisme de contrôle intervenant dès l'étape afférente sous-corticale lors de l'attention. Certaines parties de la formation réticulée mésencéphalique jouent certainement un rôle important au sein de ce mécanisme de contrôle.
On sait, en effet, que la stimulation de la formation réticulée est capable de contrôler la transmission de messages sensoriels afférents musculaires (11), somesthésiques (12), visuels (10-14) ou auditifs (18). L'absence de blocage observée dans notre observation 1, où une lésion du tegmentum mésencéphalique est très probable, confirme ces données.
Cette mise en jeu des formations centrales responsables du blocage pourrait se faire à partir du cortex. La stimulation d'aires corticales en relation étroite avec la formation réticulée peut en effet "bloquer" certaines afférences au niveau des premiers relais spécifiques (17).
D'une façon générale, les résultats obtenus démontrent le rôle important joué par le cortex lors du processus d'attention puisque les messages importants lui parviennent avec leur amplitude maximale. Ils évoquent également la mise en jeu des formations multisynaptiques du trône cérébral clans le processus concomitant de blocage des autres modalités sensorielles (fig. 11).
Ce couplage fonctionnel de l'écorce et des formations réticulaires centrales paraît ainsi être une (les conditions nécessaires de l'attention, et par là de la prise de conscience d'un signal.
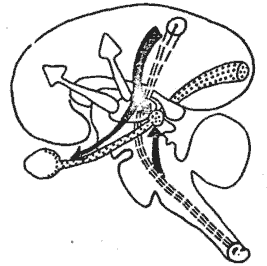
Fig. 11. - Schéma de mécanismes hypothétiques entrant en jeu au cours de l'attention. Le stimulus attractif somesthésique met enjeu des mécanismes inhibiteurs (flèches noires) à point de départ sous-cortical ou cortical, agissant sur les afférences visuelles (pointillé) soit au niveau rétinien, soit au relais géniculé. Il met en jeu également un mécanisme dynamogénique qui active l'écorce cérébrale (flèche blanche).