| Le rêve : Histoire naturelle |
| Par Michel Jouvet Médecine et Hygiène 35 : 1203-1242 |
Texte d'une conférence faite le 28 octobre 1976 à l'occasion du centenaire de la Faculté de médecine de Genève.
Le rêve, cette activité cérébrale incontrôlée a laquelle l'homme passe près de 10% de sa vie n'a pas cesse d'intriguer les physiologistes. Michel Jouvet, lors d'une conférence faite a l'occasion du 100e anniversaire de la Faculté de médecine de Genève, le 28 octobre 1976, a montré pourquoi le rêve ne peut apparaître qu'après un stade de sommeil profond, témoin électrophysiologique et biochimique de la sécurité de l'animal, car c'est seulement lorsque l'animal ou l'homme se sent en sécurité qu'il peut s'abandonner a la paralysie aveugle et sourde du rêve. On ne voit pas en effet comment l'évolution aurait pu inventer un autre moment que le sommeil pour insérer les périodes de rêve dans le continuum éveil-sommeil. Mais il reste encore bien des secrets que l'onirologie moderne essaie de dévoiler.
Je voudrais essayer, au cours de cette conférence, de vous résumer les nombreux problèmes que pose au neurophysiologiste l'activité onirique en vous exposant les résultats obtenus depuis 20 ans sur les différents fronts de recherche qui se sont attaqués au problème du rêve:
1. L'approche subjective et psychophysiologique du rêve chez l'homme a d'abord eu le mérite de situer dans le temps le rêve au cours d'une nuit de sommeil en traçant ses frontières polygraphiques.
2. L'approche objective de la neurophysiologie expérimentale a employé, de son côté, deux méthodes principales:
a.) La méthode réductioniste qui a permis d'une part de délimiter les structures responsables du rêve et, d'autre part, de découvrir quelques mécanismes biochimiques qui relient étroitement le rêve au sommeil qui en ouvre les portes.
b) L'approche historique, c'est-à-dire l'histoire naturelle du rêve : quand apparaît l'activité onirique au cours de l'évolution phylogénétique ou ontogénétique?
L'ensemble de ces approches ne permet pas cependant de répondre de façon satisfaisante à la question primordiale: qu'elle est ou quelles sont les fonctions du rêve.
I. Les limites temporelles du rêve
Il nous apparaît naturel que notre cerveau passe alternativement de l'état d'éveil à l'état de sommeil. A quel moment situer alors le rêve au cours du sommeil? A la fin du siècle dernier, Maury, professeur au Collège de France, renversa la conception idéaliste d'une âme perpétuellement éveillée au cours du sommeil en démontrant que le rêve était un phénomène épisodique ou aléatoire. Peu de réveils au cours du sommeil sont en effet suivis d'un souvenir précis de rêve. Maury émit alors l'hypothèse que le rêve apparaissait lors d'un stade intermédiaire entre l'éveil et le sommeil, soit lors de l'endormissement et avant le réveil, soit lorsque le sommeil est allégé par une stimulation externe ou interne. Cette conception devait imprégner les premiers expérimentateurs du rêve avec des techniques polygraphiques:
Entre 1953 et 1958, les travaux de l'Ecole de Chicago, avec Dement et Kleitman, démontrèrent l'existence de périodes de mouvements oculaires rapides au cours du sommeil (5). Ces périodes, d'une durée moyenne de 20 mn, surviennent environ toutes les 90 mn. Elles s'accompagnent d'un tracé électroencéphalographique similaire au sommeil léger de l'endormissement (Descending stage one). Le réveil des sujets à ce moment permet d'obtenir avec beaucoup de précision des souvenirs de rêve alors que ceux-ci sont moins fréquents et beaucoup plus estompés lorsque le réveil a lieu en dehors de ces phases. Ainsi le rêve fit son entrée en psychophysiologie. D'épisodique (ou aléatoire), le rêve devenait un phénomène rythmique doué d'une périodicité ultradienne. Pour l'Ecole de Chicago, cependant, le rêve survenait au cours d'un stade de sommeil léger. Ce concept de stade semi vigil s'exprime alors dans la terminologie de l'époque: ascending ou emerging stage one. En fait grâce aux travaux de neurophysiologie animale que devait découvrir cet état (que nous avons baptisé sommeil paradoxal), chez le chat (13), il apparu que le rêve n'était pas un stade de sommeil léger (4), mais bien un nouvel état de fonctionnement du cerveau, aussi différent du sommeil que celui-ci l'est de l'éveil (tableau). La découverte de ce nouvel état fonctionnel du cerveau devait nous entraîner vers deux voies de recherche, celle réductioniste et synchronique de la neurophysiologie expérimentale et celle holistique et diachronique.
II. Neurophysiologie expérimentale (6) à (9)
Sur le plan phénoménologique, le sommeil paradoxal (SP), qui survient par périodes de 6 mi toutes les 25 mn au cours du sommeil, s'accompagne d'une élévation importante du seuil d'éveil par rapport au stade de sommeil profond qui le précède. Il est devenu classique d'y distingue deux catégories de phénomènes toniques et phasiques. Les phénomènes toniques sont caractérisé par une activité électrique corticale rapide (similaire à celle de l'éveil) et par l'abolition totale du tonus musculaire. Les phénomènes phasiques centraux sont représentés par une activité de pointes de haut voltage au niveau du pont, du noyau genouillé latéral et du cortex occipItal. d'où le terme d'activité PGO qui lui a été donné). Cette activité est responsable des phénomènes phasiques périphériques constitués avant tout par des mouvements oculaires rapides et accessoirement de petits mouvements fins des extrémités.
Il apparaît ainsi que de nombreux systèmes de neurones sont mis en jeu de façon simultanée au cours du SP. Afin d'essayer de délimiter l'organisation de ces systèmes, notre première tactique fut d'essayer de réduire le phénomène complexe du SP à un phénomène plus simple. Malheureusement, il n'était pas question de "simplifier" le sujet d'expérience. Ni l'aplysie, ni l'escargot, ni la grenouille ne présentent de SP, tout au moins avec les critères que nous reconnaissons chez les mammifères. C'est pourquoi il nous a fallu effectuer cette simplification sur l'animal de choix des chercheurs du sommeil : le chat, en réalisant des préparations décortiquées ou porteuses de diverses sections du tronc cérébral. Je reconnais bien volontiers les limites d'une telle approche : ainsi le SP périodique de cet animal machine qu'est le chat pontique est assez loin de ressembler à celui de l'animal intact, mais au moins il nous permet de savoir que des structures responsables du SP persistent chez une préparation démunie de télédiencéphale ou de mésencéphale. Ce résultat démontre également que le SP n'est pas une propriété intrinsèque de n'importe quelle cellule cérébrale, mais qu'il dépend de certains systèmes de neurones pontiques.
Le bilan d'une vingtaine d'années de recherches sur les mécanismes du SP peut être résumé de la façon suivante: très schématiquement, le SP apparaît comme un état d'excitation cérébrale intense, au cours de laquelle la majorité des neurones semble obéir à un générateur pontique. L'excitation cérébrale se traduit sur le plan unitaire ou multiunitaire par l'augmentation rythmique de l'activité des neurones corticaux et sous-corticaux, pyramidaux ou extrapyramidaux. C'est là un fait essentiel : le faisceau pyramidal est envahi par de nombreuses décharges descendantes qui gagnent les motoneurones spinaux. Il en est de même de nombreux relais extrapyramidaux : noyaux rouges, noyaux vestibulaires, formation réticulée mésencéphalique, etc... La nature de cette excitation endogène pose de nombreux problèmes dont les principaux me paraissent être les suivants :
- Quels sont ses rapports avec les générateurs pontiques?
- Comment le cerveau reçoit-il cette information et seulement cette information, ce qui pose le problème de l'inhibition des autres signaux afférents.
- Comment le cerveau bloque-t-il les messages efférents moteurs mis en jeu par le générateur?
- Le générateur pontique est-il responsable d'un bruit aléatoire ou délivre-t-il une information?
- Enfin quels sont les mécanismes préparatoires et protecteurs qui permettent au système nerveux central de se mettre dans des conditions aussi téléologiquement dangereuses?
(A) Bien que la nature du neurotransmetteur mis en jeu par le générateur reste indéterminée, nous savons qu'au niveau de la partie dorsolatérale du tegmentum pontique, dans la région des noyaux réticularis pontis caudalis, oralis et subcoeruleus, se situe un groupe de cellules responsables de l'activité PGO.
Cette activité a le mérite d'être très individualisée au niveau du système oculomoteur et du système visuel central du chat, et son organisation anatomique a été tracée dans ses grandes lignes : en bref, de la formation réticulée pontique monte un système de neurones de très petit calibre qui croisent au niveau de la commissure supraoptique pour gagner les noyaux géniculés latéraux et le cortex occipItal. controlatéral. Le fonctionnement des générateurs est donc repéré par des informations recueillies au niveau des noyaux genouillés latéraux et des noyaux moteurs intrinsèques des globes oculaires. Un problème principal reste à résoudre : cette information privilégiée qui est recueillie au niveau du système visuel et oculomoteur est-elle seulement localisée à cet endroit, ou au contraire, n'est - elle qu'un aspect électrophysiologique privilégié d'une excitation endogène qui pourrait être recueillie dans d'autres systèmes?
Il semble que l'on puisse répondre affirmativement à la seconde question car il existe d'étroites corrélations entre la modulation de l'activité unitaire corticale ou sous-corticale en dehors du système visuel et l'activité PGO. Cette corrélation peut prendre plusieurs aspects: facilitation ou inhibition des décharges unitaires. En outre, líinactivation réversible par cryode du pédicule d'origine des PGO au niveau de l'isthme permet de supprimer la modulation de l'activité unitaire extravisuelle. Ainsi, on peut admettre qu'un très grand nombre de neurones cérébraux, appartenant aussi bien au système visuel qu'à d'autres systèmes intégratifs et moteurs sont sous la dépendance directe ou indirecte des générateurs pontiques. Cette dépendance étroite est grandement aidée par le blocage de la majorité des signaux du milieu extérieur.
(B) Inhibition présynaptique au niveau des différents relais. - Cette inhibition a été mise en évidence au cours du SP à différents niveaux: noyaux cuneatus, geniculé latéral, noyau cochléaire, etc. Le blocage des messages afférents permet donc à un grand nombre de neurones de ne plus être soumis à l'arrivée de volées afférentes extrinsèques, ce qui permet sans doute à des processus intégratifs de mieux s'établir sous l'influence des générateurs pontiques. Mais cette inhibition ne va pas sans une contrepartie téléologiquement dangereuse, c'est-à-dire l'élévation importante du seuil d'éveil.
(C) A ce blocage des afférences, s'ajoute nécessairement celui des efférences. Nous rêvons que nous courons ou que nous volons mais notre corps reste inerte car il existe une inhibition totale du tonus musculaire. Parfois peuvent apparaître, surtout chez le chien, des mouvements des pattes mais l'atonie musculaire ne permet jamais l'apparition de mouvements de redressement. Si les mécanismes intimes du blocage des efférences sont encore mal connus, nous connaissons cependant la topographie exacte des neurones responsables du blocage des efférences motrices au niveau du complexe du locus coeruleus (les efférences végétatives continuant à s'extérioriser comme en témoignent les variations respiratoires, cardiaques et tensionnelles, et peut-être l'érection dont le mécanisme reste mystérieux). Il est donc devenu possible, en détruisant ces neurones par des coagulations limitées de supprimer sélectivement l'inhibition du tonus musculaire au cours du SP. Ainsi peuvent se dévoiler des comportements stéréotypés d'orientation, d'affût, d'exploration visuelle, d'agression, de poursuite ou de défense. Ces comportements ressemblent à s'y méprendre à un comportement d'éveil. Cependant, le syndrome oculaire (myosis intense et rétraction des membranes nictitantes) est celui d'un sommeil profond. En outre, l'animal ne réagit pas aux stimulations du milieu extérieur, en particulier aux stimulations visuelles. Enfin l'activité PGO spécifique du SP que l'on observe au cours de ces épisodes est la preuve électrique que l'animal n'est pas éveillé, mais qu'il extériorise au contraire les nombreux comportements " oniriques " stéréotypés qui normalement sont bloqués par l'inhibition du tonus musculaire.
(D) L'analyse " éthologique " de ces comportements " oniriques ". - en révèle la richesse et l'organisation : ainsi, le redressement de la tête est suivi par des mouvements d'exploration de droite à gauche, puis par l'attitude d'affût typique du chat suivie parfois par la poursuite d'un objet imaginaire. Les mouvements de l'animal sont précédés chaque fois par une activité PGO. Il existe ainsi au niveau central un véritable codage, assurément fort complexe, qui est responsable des différents répertoires de comportement " onirique ". Ces comportements sont-ils acquis ou innés? Quels sont les mécanismes intimes de transduction entre les phénomènes centraux du SP, en particulier l'activité du générateur pontique, et la réalisation de ces comportements stéréotypés? Telles sont les questions essentielles que nous étudierons plus loin. Mais nous pouvons déjà répondre à la question que nous posions au début:
le générateur pontique est-il responsable d'un bruit aléatoire ou délivre-t-il une information?
On doit bien admettre que le répertoire varié des comportements " oniriques " qui survient au cours du SP en étroite liaison avec l'activité PGO centrale ne peut résulter d'un bruit aléatoire. C'est pourquoi il nous faut conclure que l'activité PGO est responsable d'une information, et sans doute d'un programme.
Il nous reste maintenant à essayer de comprendre comment un tel système de programmation peut survenir, comment il est protégé.
(E) Les mécanismes préparatoires du rêve.- le sommeil (9), (10). - L'irruption brutale du rêve au cours de l'éveil est pathologique: chez l'homme, c'est la maladie de Gélineau (narcolepsie-cataplexie) avec son effondrement dû à la perte soudaine de tonus et perte du contact avec la réalité suivi de souvenirs oniriques. Normalement, en effet, c'est le sommeil qui prépare le rêve. Certaines de ces étapes préparatoires commencent à être connues sur les plans structural et biochimique. La première étape est liée à l'endormissement. Elle s'accompagne d'une inactivation du système d'éveil mésencéphalique et d'une diminution du métabolisme des neurones catécholaminergiques. Ce phénomène ne peut se produire que si le système d'éveil n'est plus sollicité par des signaux annonciateurs de dangers soit internes (douleurs), soit externes (absence du signal olfactif ou auditif d'un prédateur, environnement non hostile ou protégé : le terrier, les arbres à sommeil des babouins, etc.).
La deuxième étape constitue le sommeil proprement dit et un cycle complet de sommeil doit, en général, précéder la première phase de SP. Des mécanismes sérotoninergiques déclenchés par le système du raphé, entrent alors en jeu. Ils contribuent encore à inactiver le système d'éveil (bien que celui-ci puisse être à tout moment activé par un signal de danger). La libération de sérotonine doit sans doute s'accompagner au niveau des effecteurs postsynaptiques des processus restaurateurs que nous attribuerons au sommeil. Parmi les messagers biochimiques qui entrent en jeu à ce moment du sommeil, certains sans doute servent à des processus de fixation synaptique, de restauration de la fatigue cérébrale. D'autres, ou les mêmes, doivent préparer les neurones à l'arrivée du rêve. Celui-ci survient lorsqu'un mécanisme de rétroaction vient mettre fin à la libération de sérotonine au niveau des projections de la partie rostrale du système du raphé. Ce signal, ainsi que l'inactivation complète de certains neurones catécholaminergiques déclenchent alors l'irruption de l'activité PGO. Les étapes finales qui permettent l'enclenchement du SP sont encore mal connues. Il semble que ce sont des neurones cholinergiques appartenant au complexe du locus coeruleus qui soient responsables de l'inhibition du tonus musculaire et peut-être de l'activité PGO.
En résumé, l'apparition du rêve semble être commandée par une série de mécanismes complexes qui s'enclenchent de façon harmonieuse. Il suffit que l'un soit absent pour que le phénomène n'apparaisse pas. Parmi ces mécanismes, il convient d'insister sur la primauté du système d'éveil dont la mise en jeu, même fugitive, est suffisante pour empêcher la mise en jeu du SP. Cette constatation nous permet de comprendre pourquoi le rêve ne peut apparaître qu'après un stade de sommeil profond, témoin électrophysiologique et biochimique de la sécurité de l'animal, car c'est seulement lorsque l'animal ou l'homme se sent en sécurité qu'il peut s'abandonner à la paralysie aveugle et sourde du rêve. On ne voit pas en fait comment l'évolution aurait pu inventer un autre moment que le sommeil pour insérer les périodes de rêve dans le continuum éveil-sommeil. S'il eut autrefois des mutants cataplectiques ou narcoleptiques, capables de s'effondrer brusquement au moment de l'attaque ou de la fuite, il faut bien admettre qu'ils ont eu peu de chance de survivre.
Les conclusions suivantes peuvent donc être tirées de l'approche réductioniste de l'activité onirique: lorsque le système nerveux n'est plus tenu en éveil par les diverses nécessités et agressions des milieux intérieur ou extérieur, une phase de sommeil préparatoire est la condition nécessaire au déclenchement du rêve. Au cours de celui-ci, des mécanismes d'origine pontique viennent isoler le système nerveux central en fermant la porte des principales afférences sensorielles et des efférences motrices (excepté la motricité oculaire). Sourd, aveugle et paralysé, le cerveau rêveur est alors prêt à recevoir des stimuli " endogènes " à partir du pont (activité PGO.). Il semble que cette activité qui envahit la plupart des structures cérébrales ne constitue pas un bruit aléatoire puisque dans certaines conditions, on arrive à montrer qu'elle est responsable de comportement moteur très complexe. Il faut donc admettre que l'activité PGO est le support d'une information ou d'un programme. Une telle information est-elle acquise (épigénétique) ou est-elle programmée génétiquement. L'approche réductioniste n'est pas encore parvenue à résoudre ce problème, mais un début de réponse a été apporté par l'étude diachronique du rêve, par son histoire naturelle.
III. L'histoire naturelle du rêve
Celle-ci peut être envisagée à la fois sur le plan phylogénétique et ontogénétique. A quel moment peut-on objectiver le sommeil paradoxal sur des bases polygraphiques certaines, chez différentes espèces animales adultes ou chez le mammifère au cours de son développement pré - ou postnatal. La réponse à cette question n'est pas si facile. Quels critères du SP doit-on privilégier? L'activation corticale pendant le sommeil comportemental? Mais qu'en est-il chez les espèces comme les poissons ou reptiles qui n'ont pas de cortex et chez qui l'activité électrique centrale ne varie pas entre les états d'activité et de quiescence? Les mouvements rapides des yeux au cours du sommeil comportemental? Que dire alors de la chouette qui ne bouge pas les yeux, mais qui présente de façon certaine les autres critères du SP? L'absence totale du tonus? Mais ce critère peut n'être pas suffisant chez les espèces dont le tonus disparaît pendant le repos ou le sommeil. Ainsi, chez l'homme, il a fallu de nombreuses explorations pour découvrir le seul groupe musculaire (houppe du menton et sous-hyoïdiens) dont l'absence d'activité tonique était spécifique du SP. Le problème est également complexe au cours de l'ontogenèse. Il faut, en effet, que les structures postsynaptiques, effectrices du SP, soient suffisamment développées pour que puissent apparaître les signes cardinaux polygraphiques du SP. Celui-ci peut très bien être présent et jouer un rôle biochimique sur les neurones sans que ceux-ci puissent exprimer, à l'échelle macroscopique où ils sont observés, une activation tonique comme cela sera le cas pour les neurones corticaux ou des brèves activations phasiques dans le système oculomoteur.
(A) Ces réserves faites, l'exploration phylogénétique a abouti au bilan suivant (12) : l'arbre généalogique du SP, exploré bien indirectement par quelques sondages chez différentes familles ou espèces, semble relativement jeune. Il n'a pas été possible, malgré de très longs enregistrements, d'observer au cours des périodes d'inactivité (sommeil?) des poissons (tanche), des amphibiens (crapaud), des reptiles (tortue, iguane, caïman) d'accident périodique accompagné de variations végétatives, de mouvements oculaires. de variations du tonus. En bref, chez ces espèces, le sommeil ou l'inactivité est monotone ou univoque. Il semble donc que les poïkilothermes (dont l'activité centrale varie peu entre éveil et sommeil) ne présentent pas de SP. Par contre, chez tous les oiseaux et mammifères étudiés jusque là, du pigeon à l'aigle, de l'opposum à l'homme, les critères cardinaux du SP sont présents et faciles à objectiver, seul l'échidné (tachyglossus aculeatus) constitue une intéressante exception puisque jusque là il a été impossible d'objectiver chez lui la présence de SP (1). Cette exception est sans doute importante car le cerveau de l'échidné possède des caractères reptiliens. Cependant, son organisation cérébrale lui permet d'avoir des caractères électriques évidents de sommeil à ondes lentes. Il semble donc bien, grâce à cette exception, que le cerveau des reptiles, même amélioré, soit incapable d'exprimer le SP. Il existe, bien sûr, des différences quantitatives considérables concernant la durée du SP et son rythme ultradien selon les espèces considérées qu'il serait trop long d'envisager ici. Quelques caractéristiques générales peuvent cependant être résumées: les espèces chassées qui ont besoin d'un temps d'éveil très long pour acquérir une nourriture faiblement énergétique à base de cellulose (herbivores, ruminants, rongeurs) ont, en général, un temps de sommeil court et la durée du SP n'excède pas 10 à 15 mn par jour, réparties en brèves périodes au cours du sommeil. Par contre, ce sont les carnivores, prédateurs, dont le sommeil est long (et protégé) qui présentent les durées de SP les plus importantes. Mais l'intrication des facteurs écoéthologiques, l'absence presque totale de données recueillies in situ dans un milieu naturel n'ont pas encore permis de repérer des rapports significatifs entre la durée du SP et d'autres caractéristiques du système nerveux central ou du comportement. Il ne semble pas y avoir, en particulier, de rapport avec les fonctions supérieures du cerveau. L'opposum et le chat ont des durées de SP plus longues que le chimpanzé ou l'homme, bien que leurs capacités d'apprentissage soient beaucoup plus limitées.
(B) Bien qu'elle procède d'une problématique différente de l'approche généalogique du SP, il convient de résumer ici les données acquises par l'étude génétique du SP qui s'est développée surtout chez la souris (3), (20). Il apparaît, en effet, que chaque souche de souris consanguines possède des caractéristiques temporelles de SP propres à la souche (durée, répartition circadienne et ultradienne). Ces caractéristiques sont transmises selon le mode mendelien. Bien plus, l'organisation temporelle des mouvements des yeux du SP possède également des caractéristiques génétiques. Ainsi la souche BALB/C a des mouvements des yeux fréquents, tandis que chez la souche C 57BR, les mouvements oculaires sont au contraire pauvres, isolés et rares. Ces caractéristiques apparaissent également liées à des caractères génétiques comme le prouve l'étude des hybrides et des croisements en retour entre la première génération et les parents des 2 souches. Si l'on se souvient qu'il existe une correspondance très étroite entre les mouvements oculaires et l'activité centrale PGO, ce renseignement est à verser au dossier de la nature de l'information qui serait délivrée par l'activité PGO au cours du SP: nous sommes ainsi en mesure d'admettre qu'il est très vraisemblable que cette information est génétiquement programmée, du moins chez la souris. La constatation de fréquences et de modes d'organisation différents des PGO chez différentes sous-espèces de papio (papio papio, papio hamadryas) va également dans ce sens (2), encore que le rôle joué par les facteurs épigénétiques n'ait pas été éliminé chez ce dernier cas. Il est sans doute prématuré d'étendre ces conclusions à l'homme et le nombre d'individus à explorer dans le cadre de la génétique des populations rend très long et difficile l'étude génétique du sommeil. Cependant, l'exploration des mouvements des yeux au cours du rêve chez trois sujets bassaris, appartenant à ce qu'il est convenu d'appeler un isolat génétique, du Sud Sénégal, in situ, et en France (dans des conditions d'environnement très différentes) nous a révélé l'existence d'une différence importante dans la répartition de leurs mouvements oculaires par rapport à un large groupe de contrôle du même âge observé à Lyon (17). L'étude de jumeaux mono et hétérozygotes devrait permettre de continuer l'approche génétique du rêve chez l'homme, mais la complexité et la diversité de groupement des mouvements oculaires chez l'homme ne rend pas facile leur classification.
(C) Si la génétique du sommeil paradoxal nous a appris qu'il pouvait exister une programmation génétique de certains aspects du SP, l'étude de l'ontogénèse du SP semble apporter des résultats similaire (14), (15), (21): très schématiquement., on peut dire que le SP est un excellent index de la dynamique des processus de maturation du cerveau. Ainsi le SP prédomine à la naissance (et constitue la presque totalité du sommeil) chez le raton et le chaton. Au fur et à mesure de la maturation postnatale, la proportion de SP décroît pour atteindre le niveau de l'adulte (soit 200 mn par jour chez le chat) au moment du sevrage qui correspond à l'achèvement de l'organisation corticale. L'augmentation considérable du SP chez les espèces nidicoles pourrait être due à des facteurs périnataux (tels que l'alimentation lactée). En fait, la démonstration qu'il s'agit bien du niveau de maturation cérébrale est apportée par l'étude des espèces nidifuges (comme le cobaye) dont la maturation cérébrale s'effectue de façon presque totale avant la naissance. Dès la naissance, la durée du SP d'un cobaye nouveau-né est, en effet, presque similaire à celle de l'adulte. Par contre, il existe une augmentation considérable du SP in utero, si bien qu'un foetus de cobaye de 45 jours (soit 20 jours avant le terme) présente des quantités de SP analogues à celles d'un raton nouveau-né (pour un index de maturation cérébrale équivalent). L'ontogénèse du SP nous apprend donc que c'est au moment où les facteurs génétiques prénataux l'emportent sur les facteurs épigénétiques que le SP est le plus important. Elle nous apprend également que la quantité de SP semble liée à la maturation et à l'organisation cérébrale. Ainsi dans le domaine de l'ontogenèse, comme dans celui de la génétique, nous retrouvons le concept de programmation et d'organisation au sein des possibles fonctions du SP.
On devrait alors s'attendre à ce que la privation de SP retentisse sur certaines fonctions cérébrale et s'extériorise au niveau du comportement. En fait, nul n'a encore apporté la preuve de l'existence d'un trouble spécifique à la suite de la privation " sélective " de SP chez l'animal adulte, car il a encore été impossible de supprimer totalement et sélectivement le SP chez l'animal nouveau-né pendant plus de quelques jours.
IV. La privation de SP
Chez l'animal adulte, il est relativement aisé de supprimer sélectivement le SP avec des méthodes instrumentales (22) (en réveillant par exemple l'animal au début de chaque épisode de SP). Ce procédé entraîne cependant une diminution relativement importante du sommeil et produit une agression non spécifique de l'organisme. Aux privations instrumentales ont ainsi succédé les privations pharmacologiques. Dans ce domaine également, il reste à prouver que les diverses drogues qui suppriment le SP n'ont pas d'autres actions sur d'autres systèmes. Enfin, l'hétérogénéité des structures qui déclenchent le SP rend illusoire une approche purement lésionnelle. Si l'on peut supprimer " sélectivement " le SP par une lésion assez extensive du tegmentum pontique, il est illusoire de penser que d'autres systèmes n'aient pas été également lésés. Cependant, le bilan des privations instrumentales est positif puisqu'elles ont démontré l'existence d'un besoin spécifique en SP. Par exemple, la privation de SP de 4 jours, chez le chat, est suivie d'une dette théorique de 200 mn x 4 - soit 800 mn de SP. Lors de la récupération qui fait suite à la privation, on assiste a une augmentation importante (un rebond) de SP qui peut atteindre jusqu'à 60% de la durée du sommeil (au lieu de 25%). Grâce à ce rebond, la " dette " de S P peut ainsi être presque entièrement remboursée. Ces notions de " besoin ", de " dette ", de " remboursement " orientent vers l'existence d'une fonction importante, mais les quelques altérations comportementales qui ont été observées chez le chat ou le rat après privation de SP: hyperexcitabilité nerveuse, diminution du seuil épileptogène, augmentation de l'agressivité, parfois diminution de l'acquisition au cours de l'apprentissage, ne peuvent pas de façon convaincante être attribuées à la privation de SP plus qu'au stress non spécifique qui l'accompagne. L'arsenal pharmacologique (qui agit sur le métabolisme des amines cérébrales) s'est modifié considérablement depuis la découverte que les inhibiteurs des monoamine- oxydases (MAO) pouvaient supprimer sélectivement le SP. Des suppressions de longue durée (semaines ou mois) du SP ont ainsi pu être obtenues aussi bien chez l'animal que chez l'homme (chez qui ces drogues sont employées à titre thérapeutique dans le cas de narcolepsie-cataplexie ou de dépression). Il n'a pas été constaté de troubles de la mémoire, ni de troubles du comportement évidents, mis à part cependant des changements notables de la personnalité et de l'humeur chez l'homme. Dans certains cas, en effet, la suppression de SP peut avoir un effet très favorable sur l'état thymique en améliorant certains états dépressifs (23).
V. Les fonctions du sommeil paradoxal
L'absence de trouble spécifique faisant suite à la privation de SP ne signifie pas obligatoirement que celui-ci ne joue aucune fonction, même chez l'adulte. C'est pourquoi l'énigme des fonctions du SP reste l'une des plus importantes de la neurobiologie. Pour terminer, nous voudrions résumer l'hypothèse que nous essayons maintenant de vérifier. selon celle-ci, le sommeil paradoxal pourrait être l'instant privilégié au cours duquel s'effectue une programmation génétique du système nerveux central (11). Cette hypothèse repose sur les constatations expérimentales exposées plus haut et que nous résumons:
1. Les mécanismes du SP apparaissent comme particulièrement adaptés aux processus intégratifs qui feraient suite à la dérépression ou à la stimulation du génome. Ainsi, les influx afférents et efférents sont bloqués par des inhibitions pré- ou post- synaptiques. Les systèmes pyramidaux et extrapyramidaux peuvent alors être mis en jeu sous l'influence d'un " pace maker " pontique sans que les mouvements segmentaires ne risquent de réveiller le rêveur.
2. Il existe des comportements stéréotypés caractéristiques de l'espèce (affût, orientation, attaque) chez le chat au cours du SP. Ces comportements peuvent être révélés après une lésion des noyaux locus coeruleus qui contrôlent le blocage des efférences. Les stéréotypes moteurs sont en liaison directe ou indirecte avec l'activité PGO centrale. On peut donc supposer que cette activité est le support d'un programme et non d'une bruit aléatoire.
3. Il existe une composante génétique dans les modalités d'occurrence des mouvements oculaires du SP (qui dépendent également de l'activité PGO). Il faut donc admettre que ce programme a une composante génétique.
4. Il existe enfin des bases génétiques qui conditionnent certains comportements.. Cela est évident chez les animaux dont les éthologistes ont recensé les nombreux comportements " innés ". Cela est très probable chez l'homme, comme en témoignent les études faites sur des jumeaux monozygotes élevés dans des milieux différents (18).
Etant donné les modifications plastiques dues à l'action de l'environnement sur les neurones cérébraux au cours de la maturation et l'apprentissage, il apparaît bien improbable que les différences héréditaires de comportement entre individus puissent être entièrement et définitivement codées par le génome lors de la maturation et l'organisation du système nerveux. On peut alors supposer qu'une programmation périodique au cours du SP, dont le codage serait à la fois temporel (par exemple le " code " des PGO) et structural (selon les circuits intéressés par cette activité), puisse être aussi efficace et adaptatrice.
Aussi efficace, car la périodicité même du SP implique une action diachronique d'une durée limitée, et donc des modifications au niveau des synapses analogues à celles qui sont utilisées au cours de certaines formes de mémorisation à court ou moyen terme.
Un tel système de programmation périodique apparaît également adaptatif. Alors qu'une programmation génétique stricte organise dans un déterminisme étroit certaines connexions nerveuses (le système rétino-tectal par exemple), une programmation périodique au cours du SP pourrait laisser beaucoup plus de liberté dans l'expression finale du programme et permettrait ainsi des possibilités d'adaptation plus larges. En effet, la programmation pourra s'effectuer au fur et à mesure des modifications historiques du système nerveux. Il apparaît ainsi possible que certains processus de mémoire à long terme (épigénétiques.) puissent être facilités ou réprimés par la programmation génétique périodique. Il existe, en effet, des preuves expérimentales montrant l'interaction à double sens du SP avec les événements historiques. Le SP peut, en effet, augmenter après certains apprentissages, tandis que la privation de SP peut entraîner soit une diminution, soit une augmentation des performances (selon la souche génétique de Souris (16, 20)).
Le problème central de cette hypothèse consiste à comprendre comment, après une phase préliminaire de sommeil dont les signaux biochimiques préparent le processus, peuvent s'effectuer au niveau de certaines cellules du tegmentum pontique, la dérépression ou la stimulation du génome et sa transcription finale en activité électrique dont le codage serait susceptible de définir l'enveloppe potentielle de certains aspects du comportement. Quelle est, d'autre part, l'étendue du message génétique qui est programmé au cours du SP? Est-ce celui du comportement spécifique de l'espèce ? Cela semble probable chez le foetus puisque le SP constitue alors la majeure partie du cycle veille sommeil. Mais on peut supposer que ce message se raccourcisse avec l'âge et il apparaît possible que cette programmation soit alors responsable de la typologie ou de la personnalité. Bien sûr, les variations individuelles de caractère ne seront pas uniquement dues à cette programmation génétique puisqu'elles pourront être également le fait de modifications historiques. Si le SP joue un rôle dans l'établissement de la typologie individuelle, il apparaît alors que l'approche statistique de la psychologie nomothétique est sans fondement. Dans cette perspective, l'étude, chez chaque -individu, des corrélations entre le " codage " du SP (tel qu'il pourrait être approché par l'analyse des mouvements oculaires.) et la " personnalité " pourrait devenir un nouveau champ de recherches, celui de l'appréciation de la part héréditaire ou innée responsable des limites potentielles de l'expression de la personnalité.
Ainsi le champ d'étude de l'activité onirique devient de plus en plus vaste. L' " onirologie " ne peut être réductionniste et oublier l'interaction du cerveau avec l'organisme et le milieu extérieur. Elle ne peut s'appuyer sur des modèles simples: ni l'aplysie, ni la grenouille ne semblent rêver au cours du sommeil. Elle ne peut s'appuyer sur les statistiques en oubliant que chaque individu est unique. Un enregistrement polygraphique de sommeil résume en quelques heures l'interaction diachronique de l'histoire récente de l'individu (ses émotions, ses acquisitions et sa fatigue au cours de l'éveil), ses bonnes ou mauvaises nuits précédentes) et le déroulement périodique d'une stimulation ou dérépression génétique qui programme son système nerveux selon un code héréditaire. Tels sont les secrets des rêves que l'onirologie moderne essaie de dévoiler.
Figure 1
 |
 |
 |
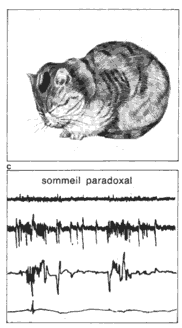 |
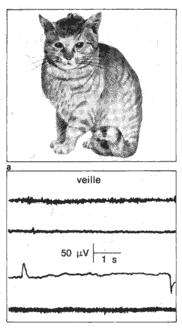
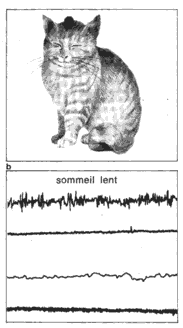
Les trois états de: éveil, sommeil lent, sommeil paradoxal. La polygraphie permet d'enregistrer chez le chat, au cours du nyctémère, bois types de tracés; Os correspondent aux trois états: veine, sommeil lent, sommeil paradoxal. EEG: électro-encéphalogramme (activité cérébrale). ** Muselles de la nuque: électromyogramme
| Tableau 1 Similitudes polygraphiques entre les périodes de réves chez l'Homme et le sommeil paradoxal du Chat |
||
| Chat | Homme | |
| Activité corticale | Rapide-similaire à l'éveil | Rapide-similaire à l'éveil ou stade 1 d'endormissement |
| Index périphériques: | ||
| Activité musculaire | Disparition activité tonique des muscles du cou | Disparition activité tonique des muscles antérieurs du cou |
| Réflexes monosynaptiques | Diminués | Diminués (H réflexes) |
| Mouvements oculaires rapides | Présents | Présents |
| Pupille | Myosis | Myosis |
| Rythme respiratoire | Apnée et irrégularité | Apnée et irrégularité |
| Aspects fonctionnels: | ||
| Seuil d'éveil | Augmenté par rapport au sommeil profond | Augmenté-égal ou supérieur au stade IV du sommeil |
| Durée | 6 mn | 20 mn |
| Périodicité | 27 mn | 90 mn |
| Effet de suppression sélective | Rebond consécutif | Rebond consécutif |
| Effets de quelques drogues | Suppression par inhibiteur monoamino-oxydase Suppression par anti-dépresseurs tricycliques |
Idem |
| Ontogenèse | Est plus important chez le nouveau-né | Idem |
| Expérience subjective | Inconnue | Rêve |
| Stricto sensu, il est évidemment impossible de prouver que le contenu subjectif du sommeil paradoxal chez le chat est l'analogue de l'activité onirique de l'Homme. Cependant, les ressemblances physiologique entre les 2 phénomènes permettent de supposer que le sommeil paradoxal est le substratum physiologique d'une fonction onirique même chez l'animal. | ||