| L'histoire naturelle du rêve |
| Conférence de Michel Jouvet |
| IMPRESSION |
| Version imprimable (Tout l'article dans une seule page) |
Le comportement onirique
Il est donc possible de prévoir ce qui doit se passer si l'on réalise une lésion bilatérale, aussi précise que possible (ce qui n'est pas si facile) du premier étage pontique du système de frein (locus coeruleus alpha) ou du faisceau descendant qui en est issu (fig.25). Le système de freinage étant détruit, rien ne pourra plus bloquer, au cours du rêve, les décharges motrices venant exciter les motoneurones alpha. Il devient ainsi possible d'observer le comportement onirique (fig. 26). Ce comportement extraordinaire peut être résumé ainsi : aucun trouble moteur, ni aucune altération du comportement ne peuvent être mis en évidence au cours de l'éveil. Le sommeil à ondes lentes est normal. Au moment où apparaissent les premières pointes PGO annonçant le sommeil paradoxal, le chat endormi ouvre les yeux et lève la tête, regardant en haut, à droite et à gauche : cette séquence d'exploration visuelle (alors que l'animal ne réagit pas aux stimuli visuels) est constante mais paradoxale : le chat tourne la tète à droite. mais regarde à gauche et inversement. Elle est ensuite suivie par des comportements variés mais imprévisibles. L'animal se lève alors brusquement et se met à marcher. Il semble courir après une proie imaginaire. s'arrêtant pour jouer avec sa proie avec le geste caractéristique du chat qui a attrapé un poisson. Brusquement peuvent survenir des comportements de peur, (avec les oreilles en arrière), de rage (avec ouverture de la gueule), ou d'attaque (avec mouvements brusques des pattes avant). Plus rarement apparaissent des mouvements de léchage. Ceux-ci ne sont pas dirigés vers un but. Parfois le chat lèche sa fourrure, plus souvent le plancher de la cage. Si l'on fixe un morceau de sparadrap sur la fourrure du chat, il la léchera pendant l'éveil. Cependant, le léchage ne sera pas dirigé à cet endroit pendant le comportement onirique. Même affamé, un chat ne se dirigera pas vers un morceau de viande qu'on lui présente pendant ces périodes. Aveugle temporairement, et sourd puisqu'il ne réagit pas aux stimulations auditives, le chat est ainsi entièrement gouverné par un système endogène qui a pris possession de son cerveau et qui le rêve. Nous n'avons observé qu'une fois un miaulement pendant ces périodes (plus de mille).
Jamais de comportement sexuel (par exemple d'attitude de rut chez des femelles en chaleur). De nombreux contrôles ont vérifié l'hypothèse que le comportement onirique représente des périodes de sommeil paradoxal sans inhibition du tonus musculaire. L'activité électrique cérébrale et le myosis sont en tous points identiques à ceux du sommeil paradoxal. Seule l'augmentation du tonus musculaire témoigne de l'absence d'inhibition du tonus musculaire. La durée moyenne des périodes de comportement onirique est de 5 minutes. Elles s'achèvent par le retour au sommeil ou par l'éveil. Les drogues qui suppriment le sommeil paradoxal (comme les inhibiteurs des monoaminoxydases) le suppriment également.
Si l'explication intrinsèque du comportement onirique est relativement facile, de nombreux problèmes restent à résoudre.
L'absence de réaction aux stimuli visuels et auditifs est compréhensible : elle témoigne du blocage complet ou incomplet des afférences visuelles et auditives mis en évidence au cours du sommeil paradoxal. Il est donc évident que ces comportements sont sans objet.
Ainsi, l'attaque onirique s'oppose à l'attaque d'une proie pendant l'éveil, ou au comportement déclenché par la stimulation de l'hypothalamus au cours duquel l'animal attaque n'importe quel objet (la main de l'expérimentateur par exemple) mais n'attaque jamais dans le vide. Le seul comportement qui pourrait offrir quelque comparaison est le jeu, lorsque les chatons peuvent courir après une feuille et parfois attaquent un ennemi imaginaire. En ce sens, comme l'avait remarqué Piaget, le rêve ressemble à un jeu intérieur du cerveau.
Les relations entre les différents comportements oniriques et l'activité PGO sont probables mais très difficiles à analyser. D'une part, la succession quasi stochastique de l'activité PGO se prête mal à toute tentative de sémantisation, d'autre part, la complexité d'un comportement de fuite ou d'attaque, mettant en jeu un nombre quasiment illimité de muscles, ne permettent pas d'analyser en détail l'activité des différents muscles en rapport avec l'activité PGO. Seule la composante d' orientation visuelle sans objet au début du comportement onirique est relativement facile à corréler avec l'alphabet PGO.
Si l'on admet la relation, au moins indirecte, entre l'activité du générateur endogène de l'activité PGO et les divers comportements oniriques, il reste encore à déterminer à quel niveau ils s'organisent, se diversifient et s'aiguillent. Sur cette question, nous n'en savons pas plus que les neurophysiologistes étudiant les divers comportements stéréotypés de l'éveil en réponse à des stimuli extérieurs ou à des stimulations centrales : importance probable de l'amygdale, de l'hypothalamus, et enfin d'une aire locomotrice située à proximité du générateur de l'activité PGO.
La dernière question est la suivante : en admettant que l'animal est aveugle et sourd, ne pourrait-il pas réagir à des hallucinations (perception sans objet) provoquées par la réponse des systèmes sensoriels à l'activité PGO ? La réponse à cette question est négative. Il est en effet possible, sur un oscilloscope, de comparer les latences entre l'arrivée de l'information PGO au niveau du nerf moteur oculaire et l'arrivée de l'information PGO dans les aires corticales visuelles. Dans tous les cas, la réponse motrice (c'est-à-dire le mouvement de l'oeil) précède de quelques millisecondes l'arrivée de l'information au niveau du cortex. Il paraît donc impossible, à moins de changer la flèche du temps, que l'effet (le mouvement oculaire de poursuite) précède la cause (c'est-à-dire l'hallucination provoquée par l'information PGO au niveau du cortex visuel). 0n est donc obligé d'admettre qu'à la fois le générateur met en jeu la poursuite oculaire et des signaux visuels endogènes : le moteur précédant le sensoriel. Alors que c'est, bien sûr, l'inverse au cours de l'éveil. Cette constatation oblige à admettre qu'il existe quelque part un système inconnu intégrant ces données avec des latences différentes. C'est là tout le problème des rapports entre les mouvements oculaires au cours du rêve et la scène onirique chez l'homme, qui est loin d'avoir été résolu.
Coupe frontale du pont chez le chat
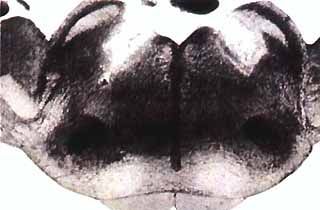
Les taches blanches bilatèrales et symetriques représentent les lésions qui interrompent le faisceau descendant issu du locus coeruleus alpha. Après de telles lésions, le animaux présentent des comportements oniriques.
Comportement onirique de peur
(qui sera suivi immédiatement après d'attaque contre une
proie imaginaire)

D'après J.P. Sastre