| Effets EEG et comportementaux des privations de sommeil paradoxal chez le chat [EEG and behavioral effects of deprivation of paradoxical sleep in cats] |
| P. Vimont-Vicary, D. Jouvet-Mounier et F. Delorme Electroencephalogr. Clin. Neurophysiol. 20 (5) pages : 439-449 (1966) |
Les premières expériences de privation de sommeil ont été réalisées en 1894 par Manaceine chez des poussins: ceux-ci mouraient après 4-6 jours sans sommeil. On doit à Légendre et Pieron (1912), puis à Schnedorf et Ivy (1939), à la suite d'expériences de privation de sommeil chez le chien, l'hypothèse de l'existence dans le milieu intérieur d' "hypnotoxines" responsables du sommeil. Depuis 1958, les études électroencéphalographiques ont permis de distinguer deux états de sommeil (Dement 1958; Jouvet et al. 1959): le sommeil lent (SL) et le sommeil paradoxal (SP) qui correspond à la phase du sommeil s'accompagnant d'activité onirique chez l'homme. Dement (1960) a montré l'existence chez l'homme d'un "besoin" de cet état de sommeil puisqu'après suppression élective il tend à réapparaître à des intervalles de plus en plus courts. Ce phénomène fut également retrouvé chez l'animal pontique chronique (Jouvet et al. 1963). Le besoin en SP peut donc s'exprimer également chez des animaux sans cortex cérébral. Enfin, des lésions électives de la formation réticulée pontique qui suppriment le SP chez le chat font apparaître des troubles du comportement qui ont été comparés à un tableau hallucinatoire (Jouvet 1962).
Le but de nos expériences a été de supprimer électivement le SP chez le chat normal pendant des durées croissantes en essayant de conserver la phase de SL, et d'étudier les effets immédiats et secondaires de cette suppression élective sur le comportement et l'EEG. D'autre part, afin d'élucider les rapports existant entre le SL et le SP nous avons été amenés à effectuer parallèlement des expériences de privation totale de sommeil.
Ce travail a fait l'objet de notes préliminaires (Jouvet-Mounier et al. 1964, 1965).
MATÉRIEL ET MÉTHODES
Après des enregistrements continus polygraphiques de contrôle (d'une durée de 24 h à 3 jours), les animaux (9 chats adultes) ont subi des privations de sommeil selon les techniques suivantes.
1. Les privations de sommeil
A. Privation de sommeil paradoxal (SP) (Fig. 1)
(1) Méthode de la piscine. Afin d'empêcher l'apparition des périodes de SP sans interrompre le sommeil de l'animal, par un stimulus trop brutal, nous l'avons placé jour et nuit au milieu d'une piscine circulaire de 2 m de diamètre, sur un socle de 10-15 cm de côté, entouré d'eau : il peut s'y accroupir et présenter des phases de sommeil avec fuseaux et ondes lentes, mais ne peut jamais parvenir au relâchement musculaire total caractéristique du SP. Dès que le tonus musculaire diminue, il se produit un déséquilibre qui réveille l'animal. Celui-ci est sorti de la piscine régulièrement deux à quatre fois par jour, pendant des durées ne dépassant pas 45 min en moyenne, où il est surveillé en permanence afin d'empêcher l'apparition du SP.
Les activités EEG, EMG et EKG des animaux sont enregistrées continuellement sur un appareil EEG pendant la privation (grâce à un déroulement lent du papier d'enregistrement de 2,5 mm / sec). Eventuellement un "onirographe" (Jouvet 1962) permet de contrôler l'activité EMG des muscles de la nuque, grâce à une camera Grass le comportement de l'animal peut être photographié à chaque minute.
Des privations de SP de 1, 2,3,4,5, 6,8, 9, 12,17,22 et 26 jours ont ainsi été effectuées chez 6 chats adultes. Nous entendons par privations de courte durée celles n'excédant pas une semaine et privations de longue durée celles qui sont supérieures à une semaine.
(2) Méthode des stimulations électriques. Des privations de SP de 48 h ont été effectuées. Les stimulations étaient appliquées (par l'intermédiaire d'électrodes cutanées fixées sur une patte postérieure de l'animal) dès l'apparition des premières pointes ponto-géniculo-occipItal.es qui annoncent le SP. Dans le cas où elles n'étaient pas suffisantes pour empêcher l'apparition du SP, ces stimulations étaient alors répétées avec une intensité croissante par l'expérimentateur qui observait continuellement les animaux.
B. Privation totale de sommeil
Elles furent effectuées chez deux animaux pendant 24 h au moyen de stimulations diverses (bruits, stimulations proprioceptives et enfin nociceptives) appliquées dès l'apparition des premiers fuseaux au niveau du cortex sensori-moteur. A trois autres reprises, nous avons obtenu une telle privation chez des animaux qui refusaient de s'asseoir sur le socle de la piscine et qui sont restés debout dans l'eau pendant des durées de 6-48 h.
2. La récupération après les privations de sommeil
Après les privations de sommeil, les animaux sont soumis à des enregistrements continus dans une cage insonorisée jusqu'à ce que leur rythme veille-sommeil soit redevenu similaire à celui observé au cours des enregistrements de contrôle. Au début de la récupération, les pourcentages de la durée des différents états de vigilance sont calculés dès que le chat s'endort. Pour faciliter la comparaison entre les pourcentages établis au cours des différentes expériences, nous les avons calculés toutes les 6 h afin d'avoir une vue plus globale de leur évolution dans le temps. Enfin, l'animal est toujours laissé au repos pendant plusieurs jours avant de subir une nouvelle expérience.
RÉSULTATS
I. Contrôles: le rythme veille-sommeil normal chez le chat
Chaque animal, après implantation chronique d'électrodes, subit des enregistrements polygraphiques dans une cage insonorisée. L'activité des différentes aires corticales (sensori-motrice, visuelle), sous-corticales (formation réticulée pontique, noyaux géniculés latéraux), l'électrooculogramme, l'activité électromyographique des muscles de la nuque et les rythmes cardiaque et respiratoire sont enregistrés de façon continue pendant 2-4 jours. Il est ainsi possible d'obtenir des résultats stables (avec un écart standard relativement faible) (Tableau I).
Les critères électroencéphalographiques sur lesquels nous nous sommes basés pour établir ces résultats sont les suivants: dans les périodes de sommeil lent (SL) nous comprenons aussi bien l'activité de fuseaux que celle d'ondes lentes (car les enregistrements à petite vitesse 0,25 cm / sec ne permettent pas de bien les distinguer); les phases de SP sont délimitées à leur début par l'apparition d'une activation corticale associée à une atonie musculaire et leur fin est annoncée par les premiers fuseaux corticaux. Une attention particulière a été portée aux phénomènes phasiques du SP, c'est-à-dire à l'activité de pointes monophasiques ponto-géniculo-occipItal.es: au niveau de la formation réticulée pontique (Jouvet et al. 1959), du noyau géniculé latéral (Mikiten et al. 1961) et du cortex visuel (Mouret et al. 1963). Ces pointes apparaissent dans les 2 min (en moyenne) qui précédent le SP. Au cours du SP elles se groupent, en majeure partie, en bouffées de 6-8/sec. Il est à noter qu'au cours du SL on peut observer quelques périodes de pointes isolées, de grande amplitude, dont la durée totale occupe 5 % ( +- 2%) de la durée d'enregistrement.
TABLEAU I
Contrôles
Durée totale d'enregistrement: 500 h. Nombre d'animaux: 9.
| E/24 h | SL/24 h | SP/24 h | ST/24 h | SP/ST | SL/ST | Durée moyenne des PSP | Intervalle moyen entre les PSP |
| 31,5% +- 7 |
52,5% +- 7 |
16% +- 2 |
68,5% +- 6 |
23% +- 4 |
77% +- 4 |
5'45'' +- 1'20'' |
27'30'' +- 1'20' |
E : éveil;
SL : sommeil lent;
SP, sommeil paradoxal;
ST : durée totale du sommeil;
PSP : phase de sommeil paradoxal;
+- : écart type.
RÉSULTATS
II. Les privations de sommeil
A. La privation de SP par la méthode de la piscine
(1) Pendant la privation:
La première semaine, l'animal adopte rapidement une position qu'il gardera par la suite: assis, puis accroupi sur le socle, il présente périodiquement des "tentatives de SP" qui entraînent une chute de la tête et un déséquilibre du corps. Les phases de SL sont fréquentes et peuvent s'accompagner alors de pointes ponto-géniculo-occipItal.es: soit isolées, soit en bouffées apparaissant par périodes de 10 sec à 1 min, elles sont interrompues par le réveil dû au déséquilibre entraîné par la diminution du tonus musculaire au cours des tentatives de SP. L'analyse EEG révèle une prédominance des fuseaux sur les ondes lentes par rapport à l'activité observée normalement pendant la phase de SL. Le taux global de fuseaux et ondes lentes corticaux est environ de 50% par rapport à la durée d'enregistrement, donc normal (Tableau I).
Au-delà de la première semaine, les tentatives de phases de SP deviennent de plus en plus fréquentes ce qui entraîne des chutes brusques et répétées de l'animal dans l'eau. Les périodes de fuseaux et ondes lentes deviennent alors plus courtes (1-2 min vers le quinzième jour, de 30 sec -1 min vers le vingt cinquième jour). Les phases de fuseaux et ondes lentes n'occupent alors plus que 30% en moyenne du temps d'enregistrement. Nous n'avons pas observé de troubles du comportement notables au cours des 8 premiers jours de privation. Le rythme cardiaque s'élève cependant de 60% en moyenne par rapport au chiffre de contrôle dès le début de la privation de SP. C'est une tachycardie sinusale régulière qui diminue légèrement pendant les périodes de SL (exemple: chat R5: contrôle: EKG pendant l'éveil, 135/min; pendant le SL, 126/min; pendant une privation de SP: EKG pendant l'éveil, 226/min; pendant le SL, 217/min).
Après une semaine, progressivement, le besoin de sommeil prédomine sur la recherche de nourriture. Après une phase de léchage très brève, les animaux, sortis de la piscine, restent souvent assis ou accroupis dans la même posture que sur le socle de la piscine et l'on assiste à des chutes répétées de la tête et parfois du corps. Ils présentent également de la photophobie et recherchent l'obscurité. Le poids a tendance à baisser, l'appétit est moyen avec une prédilection pour les liquides. Enfin, l'hypotonie musculaire est de règle et elle évoque parfois la myasthénie après de longues privations de SP (les animaux ne peuvent alors plus sauter sur une chaise pour y chercher leur nourriture). Il n'a jamais été remarqué de signes comportementaux comparables à ceux qui évoquent un tableau hallucinatoire chez le chat porteur de lésions pontiques. Chez deux chats mâles adultes enfin il fut noté des troubles du comportement sexuel à type d'excitation: tentatives répétées de coït avec des chatons mâles.
(2) La récupération.
(a) Effets immédiats. au cours des 6 premières heures: le comportement de sommeil d'un animal qui "récupère" après une privation de SP est d'autant plus caractéristique que celle-ci a été plus longue.
Après une période de léchage d'une durée variable (plusieurs heures pour les privations courtes et quelques minutes pour les privations longues) l'animal s'endort. Une phase de sommeil lent, d'autant plus courte que la privation a été plus longue (20 sec -2 min pour des privations supérieures à 8 jours), précède toujours la première phase de SP. Après de longues privations celle-ci est caricaturale et s'accompagne d'une recrudescence des phénomènes moteurs phasiques : secousses brusques et généralisées de tous les muscles du corps, caractère explosif des mouvements des yeux associés à des secousses de la tête et des mouvements rapides des vibrisses. Lorsque le SP est déclenché, il est très difficile à interrompre; après de longues privations, il n'est pas rare de le voir se poursuivre malgré des stimuli nociceptifs: s'il y a réveil, il est très court et l'animal retombe dans le sommeil paradoxal sans phase de sommeil lent intermédiaire. Les mouvements des yeux au cours des premières phases de SP sont nombreux. Cependant après les longues privations, ils sont notablement modifiés: les bouffées de mouvements des yeux apparaissent alors groupées toutes les 10 ou 20 sec, parfois simultanément aux secousses du corps, et dans leur intervalle les mouvements isolés des yeux sont plus rares qu'au cours du SP normal.
Au point de vue EEG, les bouffées de pointes ponto-géniculo-occipItal.es sont relativement plus nombreuses qu'au cours du SP normal et les pointes sont plus nombreuses dans ces bouffées que normalement. Il y a enfin une augmentation du pourcentage des pointes ponto-géniculooccipItal.es isolées au cours du SL.
Périodicité des différentes phases du sommeil. Au cours des 6 premières heures de la récupération, le pourcentage de SP est considérablement augmenté. Pour des privations inférieures ou égales à 4 jours, cette augmentation est essentiellement due à une prolongation de la durée moyenne du SP (Tableau II). Pour des privations supérieures à 4 jours, le pourcentage de SP atteint un maximum de 60% (+5%). Ce taux est constant même après des privations de très longue durée (Fig. 2) et semble donc constituer une limite. La durée moyenne du SP est alors moins augmentée qu'après des privations de courte durée, tandis que l'intervalle entre les phases de SP est beaucoup plus court (Tableau III).
(b) Effets secondaires. Pendant les heures et les jours suivants, l'aspect comportemental du SP redevient normal et les phases d'éveil sont plus fréquentes. La durée du retour à un rythme de sommeil normal est proportionnelle au temps de privation (Fig. 3). Dans nos conditions expérimentales elle est égale à la moitié du temps de privation. Cette période secondaire de la récupération est ainsi marquée par la persistance d'une fréquence accrue des phases de SP, dont la durée moyenne très augmentée au cours des premières heures redevient normale en 24 h en moyenne. Par contre, I'intervalle entre les phases de SP revient à son chiffre normal en un temps qui est fonction du temps de privation (Tableaux II et III).
Malgré cette élévation du taux de SP au cours de la récupération, il n'y a jamais de restitution intégrale de la perte totale ou "dette" en SP accumulée au cours de la privation. Si l'on considère qu'un animal présente en moyenne 4 h de SP par jour, la privation de SP entrame une dette (A) égale à 4 h multipliées par le nombre de jours de privation. Or, au cours de la récupération, le pourcentage de SP observé (B) comprend le pourcentage normal moyen quotidien (C) augmenté de celui qui représente la récupération réelle. Le pourcentage de récupération de la dette en SP qui peut s'exprimer ainsi:
((B - C) x 100) : A
est compris entre 20 et 30° pour des privations de courte durée (entre 2 et 5 jours) et s'abaisse à 10% pour des privations plus longues (entre 11 et 22 jours).
Évolution du rythme cardiaque (Fig. 4 et 5). Au cours de la récupération du SP on assiste à un ralentissement progressif du rythme cardiaque dont l'élévation était persistante pendant toute la durée de la privation de SP. La fréquence cardiaque retrouve ensuite sa valeur normale en même temps que les pourcentages des différentes phases de sommeil redeviennent normaux, c'est-à-dire en une durée moyenne égale à la moitié du temps de privation. Au cours de la récupération, les variations relatives de la fréquence cardiaque pendant le SP sont similaires à celles qui s'observent dans les conditions de contrôle.
(c) Sur un cas de mort après privation de SP de 26 jours. Cet animal étant mort à la suite de cette expérience, il nous semble intéressant de décrire ce cas unique, car nous n'avons plus voulu poursuivre ensuite de privation de SP aussi longue par cette méthode chez d'autres animaux. Cet animal avait déjà subi des expériences de privation de SP de 2, 4, 5 et 9 jours au cours des 6 mois antérieurs, à des intervalles espacés.
Jusqu'au 20ème jour de privation de SP, l'état général et le comportement de l'animal étaient comparables à ceux observés au cours de privations de longues durées chez les autres animaux. C'est entre le 20ème et le 26ème jour que l'état général s'est aggravé, avec formation d'abcès localisés des soles plantaires et amaigrissement (600 g sur 3200 g de poids initial) qui ont motivé l'arrêt de l'expérience. A cette date, les "tentatives de SP" étaient très rapprochées (toutes les 2 min) et le pourcentage de fuseaux et ondes lentes de plus en plus bas (10%).
La récupération qui débuta au milieu du 26ème jour s'avéra être très anormale: le premier jour, l'animal dont le rythme cardiaque était de 200/min présentait une phase de SP 20 sec après son installation dans la cage d'enregistrement. Au point de vue comportemental et polygraphique, les signes de récupération étaient comparables à ceux observés après de longues privations de SP. Par contre, de courtes phases de SP se succédèrent à une fréquence beaucoup moins élevée si bien que le pourcentage de SP ne dépassa pas 36% (au lieu de 60%) au cours des 6 premières heures.
D'autre part, vers la troisième heure de récupération, survint un accès de polypnée intense à 135/min, qui persista pendant 1 h et céda au début d'une phase de SP. Enfin, les périodes de fuseaux et d'ondes lentes s'accompagnaient d'une atonie musculaire permanente et, pendant l'éveil, l'animal était hypotonique au point de ne pas pouvoir rester assis. Cet état associé à un comportement léthargique persista pendant les 3 premiers jours.
Au troisième jour, le taux de SP (sur 24 h) était anormalement bas (10%) et le rythme cardiaque toujours élevé (200/min). En dehors des phases de SP on notait en outre l'apparition d'épisodes atypiques: d'une part, il survenait, après des périodes de sommeil lent, des phases d'activité corticale rapide, d'une durée de 3-5 min, associée a une atonie musculaire, mais sans mouvements des yeux, pendant lesquelles le comportement était celui du sommeil. D'autre part, immédiatement après des périodes d'éveil typique et sans phases de SL préalables apparaissaient des phases d'activation corticale avec atonie musculaire et mouvements des yeux qui ressemblaient en tous points au SP et évoquaient les attaques de narcolepsie telles qu'elles ont été décrites chez l'homme (Rechtschaffen et al. 1963) (Fig. 6).
Au cinquième jour, l'état général de l'animal semblait s'améliorer et l'hypotonie d'éveil avait disparu. Par contre, le rythme cardiaque devenait irrégulier, soit brady- soit tachyarythmique et l'animal mourait par fibrillation ventriculaire. L'autopsie a révélé des hémorragies au niveau des glandes surrénales et un encéphale macroscopiquement normal.
TABLEAU II
Évolution de la durée moyenne, de l'intervalle moyen et du pourcentage du SP par 24 h pendant les 9 jours de récupération (R) après une privation de SP de 22 jours chez un animal
| Durée moyenne du SP | Intervalle moyen | % SP/24 h | |
| Contrôle | 6'23" | 27'14" | 19,4 |
| R. 1° jour | 9' | 8'40" | 48 |
| R. 2° jour | 6'06" | 12' | 31,5 |
| R. 3° jour | 6'50" | 17' | 27,9 |
| R. 4° jour | 6'10" | 19' | 24,1 |
| R. 5° jour | 7' | 23' | 22,3 |
| R. 6° jour | 6'20" | 22' | 22 |
| R. 7° jour | 6'30" | 22' | 20,6 |
| R 8" jour | 5'29" | 25' | 20 |
| R. 9° jour | 5'20" | 26' | 19 |
TABLEAU III
Moyennes (calculées par périodes de 6 h) des durées du SP et des intervalles entre les phases de SP après des privations de SP de 5 et 22 jours au cours des 48 premières heures de récupération (R) chez 2 animaux
| Durée privation de SP | Heures et jours de R | 0-6h | 6-12h | 12-18h | 18-24h | 24-30h | 30-36 h | 36-42 h | 42-48 h |
| 5 jours | Durée moyenne (normale 6'30") | 18' | 9' | 11' | 8' | 7' | 6'20" | 7' | 5'45" |
| 5 jours | Intervalle moyen (normal 23') | 9' | 18' | 18' | 23' | 18' | 18'30" | 20'10" | 21' |
| 22 jours | Durée moyenne (normale 6'23") | 10'20" | 9'50" | 5'30" | 7' | 4'30" | 6' | 8' | 5'30" |
| 22 jours | Intervalle moyen (normal 27'14") | 7'06" | 8' | 7'30" | 12'30" | 8' | 12'30" | 13' | 18'55" |
RÉSULTATS
II. Les privations de sommeil
B. Privation de SP par les chocs électriques
Pendant la privation, le nombre et l'intensité des stimulations nécessaires pour supprimer l'apparition du SP augmentent progressivement: l'intervalle entre les stimulations s'abaisse alors à un chiffre moyen minimum de 2'30" dès la 18ème h (Fig. 7).
Au cours de la récupération, les animaux paraissent plus anxieux qu'après des privations de durée identique réalisées par la méthode de la piscine. Aussi, le taux maximum de récupération en SP n'a-t-il lieu qu'entre la 6ème et la 12ème h car au cours des 6 premières heures les chats présentent encore de longues phases d'éveil entre les périodes de SP comme si une certaine forme de conditionnement par les chocs était apparue. L'élévation du taux de SP pendant les 24 premières heures de la récupération est cependant identique à celle observée après privation de SP par la méthode de la piscine.
RÉSULTATS
II. Les privations de sommeil
C. La privation totale de sommeil
Il est apparu qu'il était extrêmement difficile d'empêcher l'apparition de fuseaux corticaux chez des animaux situés dans des conditions normales d'enregistrement. Les différents stimuli, efficaces pendant les premières heures de la privation de sommeil (bruits, déplacement de la cage ou de l'animal) répétés toutes les 30 sec deviennent rapidement incapables d'empêcher l'apparition de l'activité de fuseaux au niveau du cortex sensori-moteur. Dès la 5ème heure et jusqu'aux dernières heures de la privation les stimuli doivent être répétés très souvent (toutes les 20 sec en moyenne) et doivent être de forte intensité et souvent même douloureux pour entraîner une activation corticale de quelques secondes. Ainsi, malgré la répétition incessante des stimuli, un taux minimum de fuseaux corticaux (3% en moyenne du temps total d'enregistrement) persiste; c'est pourquoi une durée de 24 h semble être difficile à dépasser avec cette méthode.
La récupération est par contre très différente de celle que l'on observe après privation sélective de SP. Il n'y a en effet pas de "rebound" immédiat de SP mais au contraire une augmentation relative du SL (Fig. 8).
DISCUSSION
A. Au cours de la privation
La technique de privation par la méthode de la piscine apparaît sélective vis-à-vis du SP. En effet, la proportion de SL n'est pas diminuée par rapport au contrôle et l'apparition de plus en plus fréquente des tentatives de SP est comparable à celle que l'on observe avec la méthode des chocs électriques. Après le 8ème jour cependant, la récurrence presque invincible du SP provoque à son tour un état de privation relative du SL. Cette technique a donc permis la mise en évidence d'un "besoin" de SP que traduit nettement l'intervalle de plus en plus court entre les tentatives de SP, ce qui confirme les résultats de Dement chez l'homme (1960). Ces résultats révèlent d'autre part que l'activité de pointes ponto-géniculo-occipItal.es constitue un des meilleurs signes précurseurs du SP qui tend à augmenter (même au cours du SL) au fur et à mesure de la privation. Ces pointes, qui dans des conditions normales accompagnent les phénomènes phasiques du SP (mouvements des yeux, des vibrisses, etc.) apparaissent ainsi comme l'expression de mécanismes différents de ceux qui sont responsables des phénomènes toniques du SP (inhibition du tonus musculaire, activation corticale). Le facteur neuro-humoral hypothétique responsable du besoin en SP parait agir au niveau du tronc cérébral inférieur (et probablement du pont): la récurrence de plus en plus rapide des tentatives de SP au cours de la suppression par chocs électriques chez le chat pontique chronique est en effet identique à celle qui apparaît chez le chat normal; ainsi aucune structure cérébrale située en avant du pont n'est nécessaire à l'apparition du besoin en SP (Jouvet et al. 1963). D'autre part, les pointes ponto-géniculo-occipItal.es dépendent également de la formation réticulée pontique puisqu'elles persistent au niveau du pont (Jouvet 1962) (et disparaissent en avant) après section du tronc cérébral à un niveau prépontique (Hobson 1965). En l'absence de données humorales ou neurohumorales, il est difficile de trouver l'explication des troubles comportementaux et végétatifs communs aux animaux privés pendant plus de 8 jours (et qui pourraient être provoqués par l'accumulation d'un agent neuro-humoral). Ce tableau se caractérise surtout par de la somnolence, de l'hypotonie musculaire, une tachycardie sinusale et un degré important d'éréthisme sexuel. Il est possible que l'hypotonie musculaire et la tachycardie soient dues à la fatigue musculaire entraînée par la position assise ou accroupie des animaux qui ne peuvent se coucher. Cependant, cette hypothèse ne permet pas d'expliquer la lenteur du retour à un rythme cardiaque normal et l'hypothèse d'une tachycardie centrale, liée à l'accumulation d'un facteur neuro-humoral au niveau du tronc cérébral inférieur au cours de la privation, est plus plausible. Elle permettrait d'expliquer alors le retour du rythme cardiaque à la normale en même temps que le taux de SP. L'excitation sexuelle pourrait traduire aussi la réaction des structures hypothalamiques ou limbiques (références dans MacLean 1962) qui commandent le comportement sexuel à l'augmentation d'un possible agent neuro-humoral. Il convient enfin de signaler l'absence de troubles hallucinatoires du type de ceux décrits par Dement (1960) chez l'homme après des privations d'activité onirique de plusieurs jours. Nous n'avons pas non plus observé de tableau pseudohallucinatoire identique à celui décrit par Jouvet (1962) après des lésions de la formation réticulée pontique qui suppriment le SP. Dans ce dernier cas cependant, les mécanismes apparaissent différents puisque les lésions de la formation réticulée pontique qui entraînent ce tableau ne suppriment que l'inhibition du tonus musculaire et entraînent une augmentation considérable d'activité de pointes ponto-géniculo-occipItal.es au cours du sommeil lent, aussi les animaux porteurs de ces lésions présentent au cours des états pseudo-hallucinatoires les phénomènes phasiques et oculaires du SP sans inhibition du tonus musculaire (Jouvet et Delorme 1965). La technique des chocs électriques ou celle de la piscine en réveillant l'animal par un stimulus douloureux ou sa chute dans l'eau ne permet pas à un tel tableau d'apparaître.
DISCUSSION
B. La récupération
L'existence d'un "rebound" de SP après privation a été notée pour la première fois par Dement (1960) chez l'homme. Elle a ensuite été confirmée par Khazan et Sawyer (1963) chez le lapin après privation de SP par un bruit intense.
Certaines caractéristiques de ce "rebound" méritent d'être brièvement discutées car elles ne semblent pas encore avoir été décrites:
La première réside dans l'exagération des phénomènes phasiques du SP (au point de vue comportemental et EEG). Il semble ainsi que l'accumulation de l'agent neuro-humoral responsable du SP agisse avant tout sur les phénomènes phasiques déclenchés au niveau du pont, et secondairement seulement sur les phénomènes toniques.
La deuxième caractéristique est constituée par le taux limite du "rebound" qui est atteint après 3 ou 4 jours de privation. Ce taux de 60% (au cours des 6 premières heures de sommeil) ne peut ensuite être dépassé, même après des privations de plusieurs semaines, et semble ainsi constituer une limite biologique. Chez l'homme ce taux limite est également de 60% puisque Dement (1965) cite des pourcentages de SP de 60,2% et de 62,2% au cours de la première nuit de récupération après des privations sélectives de SP de 15 et de 16 jours. L'explication de cette limite pourrait être donnée par la troisième caractéristique du "rebound" qui est son caractère périodique nécessaire.
En effet, quelle que soit la longueur de la privation, jamais la récupération ne s'effectue de façon continue en une seule phase prolongée de SP. Au contraire, I'augmentation de la durée moyenne du SP (qui se voit surtout après les privations courtes) ne persiste que quelques heures et c'est avant tout aux dépens de la durée des intervalles que s'effectue le "rebound". La nécessité de l'intervention de périodes de SL avant chaque phase de SP pourrait alors traduire l'existence d'une nouvelle synthèse de facteurs de type enzymatique nécessaire au métabolisme de l'agent neuro-humoral accumulé au cours de la privation de SP.
Le quatrième aspect de la récupération consiste enfin dans son caractère durable (sa durée est égale à la moitié de celle de la privation). Si l'on admet l'hypothèse de l'accumulation d'un facteur de type neuro-humoral pendant la privation, il faudrait alors penser que celui-ci se trouve d'abord stocké sous une "forme liée" avant de pouvoir être métabolisé peu à peu sous une forme libre. On est alors enclin à admettre qu'il pourrait exister un rapport entre la forme liée et la tachycardie d'origine centrale. Cette constatation oriente vers la série des catécholamines, ce qui n'est pas en contradiction avec la démonstration du rôle favorisant de la DOPA dans l'apparition du SP après réserpine (Matsumoto et Jouvet 1964).
DISCUSSION
C. Rapport entre SP et SL
L'absence de "rebound" de SP après privation totale de sommeil confirme les résultats obtenus chez l'homme par Williams et al. (1962), mais est en contradiction avec ceux de Kiyono et al. (1965): la technique de privation employée par ces derniers auteurs (sans contrôle EEG) doit sans doute être incriminée car il est possible que des périodes de SL à expression électroencéphalographique aient pu survenir malgré un comportement d'éveil (analogues aux épisodes de "microsleep" observés chez l'homme) (Williams et al. 1962).Il apparaît ainsi que la récupération d'une privation totale de sommeil s'effectue d'abord aux dépens du SL et ensuite seulement aux dépens du SP. Le SL paraît donc comme une étape préliminaire habituelle à l'apparition du SP (ce qui explique le caractère périodique de la récupération.)
Il convient cependant de remarquer que la précession du SL n'est pas un facteur obligatoire de l'apparition du SP puisque dans des conditions pathologiques (qui associent privation de SL et SP comme dans le cas ayant entraîné la mort de l'animal), on a pu observer des transitions directes entre l'éveil et le SP. Si cette éventualité est rare, elle n'en révèle pas moins que le SP peut survenir dans certaines conditions sans phase de SL préalable. Cette "narcolepsie expérimentale" ressemble à la transition directe éveil-SP du chaton nouveau-né (Valatx et al. 1964) et aux cas de narcolepsie décrits chez l'homme par Rechtschaffen et al. (1963).
Résumé
1. Des privations sélectives de sommeil paradoxal (SP) ont été effectuées sous contrôle EEG chez des chats chroniquement implantés pendant des durées de 1 à 26 jours selon différentes méthodes (piscine, chocs électriques). Des privations de sommeil total ont également été effectuées pendant 24-48 h.
2. Au cours de la privation sélective de SP, on assiste à l'apparition d'un "besoin en SP" caractérisé par la diminution progressive des intervalles entre les tentatives de SP.
3. Après des privations de SP supérieures à une semaine quelques troubles discrets du comportement apparaissent. lls sont caractérisés par de la somnolence, de l'hypotonie musculaire et de l'excitation sexuelle. ll existe également une tachycardie importante. Il n'a pas été vu de tableau ressemblant à des hallucinations.
4. Après privation sélective de SP il y a, au cours du sommeil de récupération, un "rebound" sélectif de SP. Les caractéristiques de ce "rebound" sont soulignées. Il s'accompagne d'une augmentation des phénomènes phasiques du SP. Il ne peut dépasser un taux limite de 60%, du temps de sommeil au cours des 6 premières heures. Il s'effectue de façon périodique pendant une durée égale à la moitié du temps de privation. Le retour à un rythme cardiaque normal suit fidèlement le retour du SP à la normale. Enfin, il n'y a jamais de récupération totale de la dette en SP accumulée au cours de la privation.
5. Un animal est mort le 5ème jour de la récupération après une privation de 26 jours de SP après avoir présenté à de nombreuses reprises un tableau de narcolepsie au cours duquel le SP faisait directement suite à l'éveil.
6. Après privation des deux états de sommeil: sommeil lent (SL) et sommeil paradoxal (SP), il n'y a pas de "rebound" en SP mais au contraire une augmentation relative du SL.
7. Ces résultats sont enfin discutés à la lumière d'une hypothèse neuro-humorale du SP.
SUMMARY
EEG AND BEHAVIOURAL EFFECTS OF DEPRIVATION OF PARADOXICAL SLEEP IN THE CAT
1. Selective deprivation of paradoxical sleep (PS) had been carried out in cats with chronically implanted electrodes for 1-26 days by different methods (swimming pool or electric shocks). Total deprivation of sleep was also carried out for 24-48 h under EEG control.
2. During selective PS deprivation, a need for PS appears, requiring an increasing number of awakenings on successive days in order to prevent it.
3. After PS deprivation longer than 8 days some discrete behavioural disturbances may appear: mostly drowsiness, muscular hypotonia and hypersexuality. There is also a constant tachycardia. Behaviour suggesting hallucinations was not seen.
4. In sleep during recovery, after selective PS deprivation, a selective rebound of PS appears. Its characteristics are emphasized: it is associated with an increase of the phasic phenomena of PS; it is always periodically interrupted by slow sleep and its duration is equal to half the duration of PS deprivation. The return to a normal heart rate closely follows the return to a normal amount of PS. Finally, there is never complete recovery of the debt of PS which is accumulated during deprivation.
5. One cat died on the 5th day of recovery, after a 26 day PS dc privation, after numerous episodes of narcolepsy during which waking was directly followed by PS, without an intermediate episode of slow sleep.
6. Deprivation of both slow and paradoxical sleep is not followed by a PS rebound but on the contrary by an increase of slow sleep.
7. These results are discussed in the light of a neuro-humoral theory of PS.
Figure 1 : Schéma de l'installation utilisée pour la privation de SP
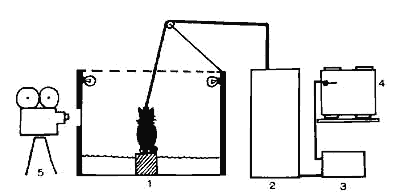
L'animal assis sur le socle dans la piscine (1) est relié à l'appareil EEG (2) sur lequel est branché un onirographe (3) inscrivant sur le cylindre enregistreur (4) I'activité électromyographique des muscles de la nuque. Une caméra (5) peut éventuellement filmer le comportement de l'animal.
Figure 2 : Existence d'un taux limite de SP au cours de la récupération
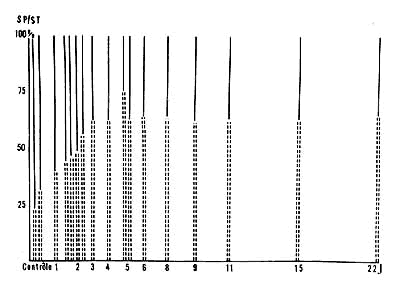
En ordonnées: pourcentage de sommeil paradoxal (SP) par rapport
au sommeil total (ST) au cours des 6 premières heures de récupération.
(Résultats obtenus au cours de 15 expériences chez 6 chats).
En batons: pourcentage de sommeil paradoxal par rapport au sommeil total.
En noir: proportion de sommeil lent par rapport au sommeil total.
En abcisses: Durée de la privation de SP en jours.
Les chiffres de contrôle ont été obtenus grâce à des enregistrements de longue durée (de 24-60 h) effectués avant les expériences de privation de SP.
Figure 3 : Courbes représentant l'évolution des pourcentages de sommeil paradoxal
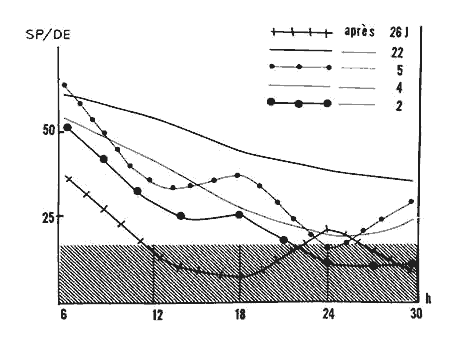
...par rapport à la durée d'enregistrement (SP/DE) pendant les 30 premières heures (h) de récupération après des privations de SP de 2, 4, 5, 22 et 26 j. Pour le cas de la privation de 26 j voir (c).
Zone hachurée: pourcentage de SP/DE de contrôle.
Figure 4 : Diagramme représentant les récupérations du rythme cardiaque
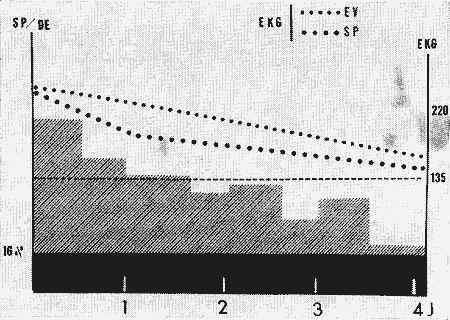
...et du taux de SP par rapport au nycthémère (SP/DE) chez un animal après une privation de SP de 8 jours.
En ordonnées:
à gauche: pourcentage de SP/DE (zone hachurée claire);
pourcentage moyen de SP/DE chez le chat normal (zone hachurée foncée);
à droite: rythme cardiaque (systoles/min) pendant l'éveil et le sommeil paradoxal (SP).
Ligne pointillée horizontale: moyenne du rythme cardiaque calculée sur 24 h.
En abscisses: jours de récupération.
DE: durée d'enregistrement,
EV: éveil.
Figure 5 : Corrélation entre la durée de récupération d'un rythme de sommeil normal
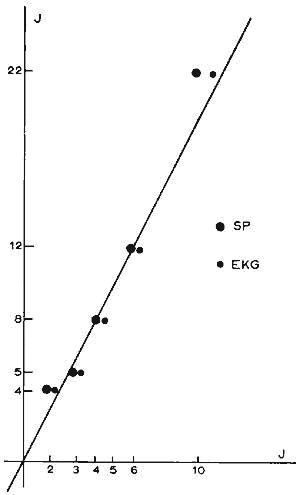
... et celle d'un rythme cardiaque normal après différents temps de privation de sommeil.
En ordonnée: temps de privation en jours (J).
En abscisse: temps de récupération (qui est égal
à la moitié du temps de privation).
Gros points: sommeil paradoxal.
Petits points: fréquence cardiaque.
Figure 6 : Narcolepsie au cours de la récupération après privation de SP pendant 26 jours
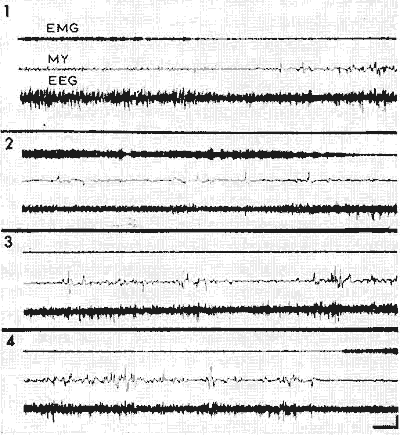
1: Contrôle: succession normale de SL et de SP (caractérisé par des mouvements des yeux (MY), l'abolition de l'EMG de la nuque (EMG) et une activité rapide du cortex visuel avec des pointes (EEG).
2, 3, 4: Au cours de la récupération après privation de SP pendant 26 j, succession pathologique de périodes d'éveil et de SP sans phase de SL intermédiaire.
Les trois tracés se font suite sans interruption, chaque ligne représente 2 min d'enregistrement. Calibrages: 6 sec et 50 microV.
Figure 7 : Evolution de la durée des intervalles séparant les tentatives de sommeil paradoxal
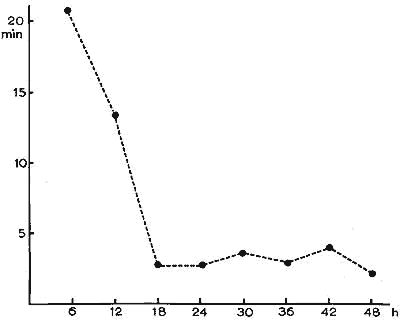
... (SP) au cours de la privation de SP par la méthode des chocs électriques.
En ordonnée: temps en minutes (moyenne par périodes de
6 h) entre les tentatives de SP.
En abscisse: durée de la privation en heures.
Figure 8 : Evolutions comparées de la récupération du rythme de sommeil
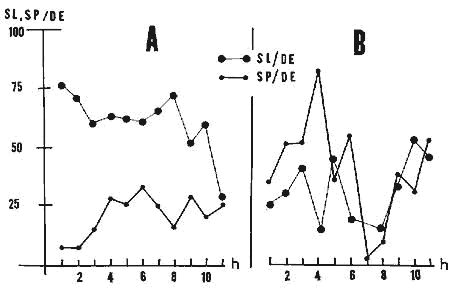
... après 48 h de privation totale de sommeil (A) et après 48 h de privation sélective de SP (B). Voir le texte.
Pour A et B:
En abscisses: heures de récupération.
En ordonnées: pourcentage de sommeil paradoxal (SP) et de sommeil
lent (SL) par rapport à la durée d'enregistrement (DE).
Gros points: SL.
Petits points: SP