| Action de la caféine sur l'activité électrique cérébrale |
| Note de M. Jouvet, 0. Benoit, A. Marsallon et J. Courjon préSentée Par J. F. Cier Extrait des Comptes rendus des séances de la Société de Biologie. Séance du 8 juillet 1957. - Tome CLI. N° 8-9. 1957, p. 1542 |
La caféine constitue l'une des substances psychotoniques les plus anciennement et universellement connues. Nous étudions ici son action sur l'activité électrique cérébrale.
Matériel et méthodes
Deux groupes de méthodes sont utilisées chez le Chat :
1) préparation encéphale et cerveau isolé (1, 2) ;
2) préparations curarisées au flaxédil (*) et destructions électives (les formations centrales méso-diencéphaliques au moyen de coagulations. L'EEG est enregistré au moyen de vis fixées sur le crâne et les structures sous-corticales au moyen d'électrodes bipolaires concentriques orientées stéréotaxiquement. Le contrôle du niveau (les sections du tronc est effectué macroscopiquement, celui des coagulations par examen microscopique.
Résultats
I. ACTION DE LA CAFEINE SUR L'ACTIVITÉ ÉLECTRIQUE CÉRÉBRALE.
Sur un tracé EEG synchronisé d'animal curarisé, l'injection intraveineuse de 1 à 3 mg/kg (la caféine entraîne l'apparition d'une activité électrique corticale et sous-corticale rapide (35 à 45 c/sec) et de bas voltage (fig. 2 A et B). A dose plus forte (40 à 50 mg/kg), la caféine fait apparaître une activité critique épileptique généralisée.
Fig. 1. - Comparaison de l'action du maxiton et de la caféine sur l'activité électrique du "cerveau isolé".
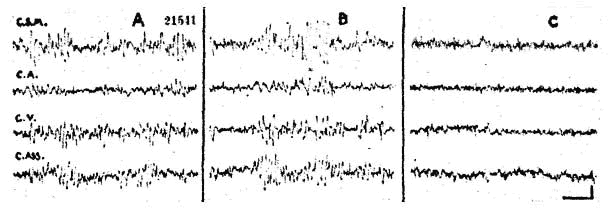
Chat. Flaxédil ; section intercolliculaire. A : activité après la section. B : 1 minute après l'injection I.V. de 5 mg/kg de maxiton : aucune activation. C : 1 minute après injection I.V. de 15 mg/kg de caféine : activation.
II. LIEU D'ACTION.
L'activité rapide et de bas voltage provoquée par l'injection de caféine s'observe également sur les préparations "encéphale isolé" et "cerveau isolé". Elle persiste également après coagulation totale des formations grises diencéphaliques (coordonnées A-11 Horsley-Clarke), tandis que l'injection de maxiton ne provoque plus aucune accélération du tracé cortical (fig. 1 . A, B et C). Enfin l'injection de caféine peut également entraîner l'activation d'un lambeau cortical isolé par des sections sous-piales.
Sur les préparations déconnectées des formations sous-corticales, la caféine déclenche une activité critique épileptique. Cette activité nécessite des doses moindres (20 à 30 mg/kg) que les préparations entières.
Fig. 2. - Action de la caféine sur les potentiels évoqués.
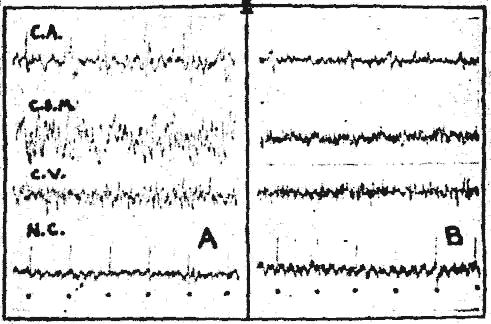
A. Chat. Flaxédil, réponses du cortex acoustique (C. A.) et du noyau cochléaire dorsal (NC) à des clics.
B. 60 secondes après l'injection de 5 mg/kg de caféine réduction importante des réponses corticales sans changement d'amplitude des réponses du noyau cochléaire
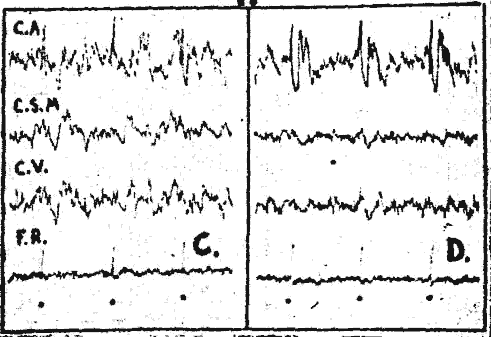
C. Chat. Flaxédil, réponses du cortex acoustique monopolaire (C. A.) et de la formation réticulée" mésencéphalique à des clics, après coagulation du thalamus en A 11.
D. 60 secondes après l'injection de 15 mg/kg de caféine : activation et augmentation d'amplitude des réponses corticales portant surtout sur la réponse lente. Noter aussi l'augmentation des réponses réticulaires.
III. ACTION DE LA CAFÉINE SUR LES RÉPONSES A DES STIMULATIONS SENSORIELLES
Les stimuli les plus généralement utilisés ont été des clics. Sur un animal curarisé au flaxédil, l'injection de caféine entraîne une réduction importante des potentiels évoqués corticaux. Ce "blocage" est analogue à celui qui s'observe lors de la réaction d'arrêt provoquée par une stimulation nociceptive ou une stimulation réticulaire. Cette réduction d'amplitude des réponses corticales ne s'accompagne d'aucune modification des réponses auditives au niveau du premier relais spécifique (noyau cochléaire dorsal) (fig. 2 : A et B).
Par contre, sur l'animal diencéphalique, chez qui les voies (le projection réticulocorticales sont détruites au niveau thalamique, l'injection de caféine entraîne, en même temps qu'une activation corticale, une augmentation d'amplitude (les réponses corticales intéressant surtout la phase lente du potentiel évoqué, Les réponses auditives de la formation réticulée mésencéphalique (A 2. L 2. H 2.) enregistrées caudalement à la lésion montrent aussi une augmentation d'amplitude (fig. 2 : C et D).
Discussion
La caféine agit donc sur le système nerveux central de façon complexe.
1. Son action au niveau de la formation réticulée mésencéphalique est objectivée par le déclenchement d'une activité rapide et l'augmentation des réponses évoquées au niveau mésencéphalique,, alors que toute influence corticifuge est éliminée par coagulation diencéphalique. Cette action semble s'exercer électivement sur le système réticulé activateur ascendant qui contrôle les réponses évoquées corticales (3, 4, 5). Par contre, les formations centrales contrôlant les afférences spécifiques (6, 7), ne sont pas mises en jeu, comme en témoigne, l'absence d'inhibition des réponses au niveau du premier relais spécifique.
2. L'action corticale directe de la caféine est signée par l'activation de la préparation diencéphalique et du cortex isolé. Cette action diffère cependant de celle d'autres substances capables d'activer l'écorce déconnectée des formations multisynaptiques centrales (physostigmine, anticholinestérase 17), car il n'y a pas diminution (les potentiels évoqués, mais augmentation. Il ne s'agirait donc pas en ce cas d'une désynchronisation corticale. L'action de la caféine sur le cortex est plutôt de type strychnique. C'est cette action strychnique qui serait également responsable des pointes obtenues par l'application directe de caféine sur le cortex (8).
3. Sur le cerveau entier, l'action de la caféine sur les formations multisynaptiques centrales tend à l'emporter sur l'action corticale directe, car le contrôle réticulaire des potentiels évoqués corticaux prédomine sur l'action facilitatrice de type strychnique. Cette mise en jeu prédominante de l'activité de contrôle réticulaire expliquerait également la diminution du seuil épileptogène à la caféine sur l'animal diencéphalique.
Ainsi la caféine, par ses deux modalités d'action, prend place entre le groupe des substances dites psychotoniques agissant électivement sur ces formations centrales du tronc cérébral [amphétamines, maxiton (9, 10, 11), rItal.ine (12), adrénaline (9, 13, 4, 5)] et celles qui agissent à un niveau cortical [anticholinestérase (16)]. Ces propriétés rendent compte de son action sur les fonctions sensorielles et intellectuelles.
(*) Triiodoéthylate gallamine (Spécia).
(1)
F. Bremer, C. R. Soc. Biol. 1935, t. 118, p. 1235.
(2) F. Breiner, C. R. Soc. Biol., 1936, t. 122, p. 460.
(3) F. Bremer et C. Terzuolo, Arch. Int. Physiol., 1954,
t. 62, p. 157.
(4) H, W . Magoun, Physiol. Rev., 1950, t. 30, p. 455.
(5) G. Moruzzi et H. W. Magoun, E. E. G. Clin. Neurophysiol.,
1949, t. 1, p. 455.
(6) K. E. Hagharth et D. 1. B. Kerr, J. Neurophysiol.,
1954, t. 17, p. 295.
(7) M. Jouvet et .1. E. Desmedt, C. R. Acad. Sc., 1956,
p. 1916.
(8) B. Libet et B. W, Gerard, J. Neurophysiol., 1939,
t. 2, p. 153.
(9) M. Bonvallet, P. Dell et G. Hiebel, C. R. Soc. Biol.,
1953, t. 157, p. 1162.
(10) M. Bonvallet, P. Dell et G. Hiebel, E. E. G. clin.
Neurophysiol. 19.54, t. 6, P. 119.
(11) G. Hiebel, M. Bonvallet, P. Huvé et P. Dell,
Sent. Hôp. Paris, 1954, t. 30, 1880.
(12) M. Jouvet, A. Marsallon et J. Courjon, Experientia
(sous presse).
(13) P. B. Bradley et J. Elkes, J. of Physiol., 1953,
t. 120, p. 14.
(14) R. W. Porter, Amer J. Physiol., 1952, t. 169, p.
629.
(15) A. B. Rothballer, E. E. G. Clin. Neurophysiol.,
1956, t. 8, p. 603.
(16) J. E. Desmedt et G. La Grutta, J. of Physiol., 1957,
t, 136, p. 20.