| Contrôle central des messages acoustiques afférents |
| Note De MM. Michel Jouvet et Jean-Édouard Desmedt, Extrait des Comptes rendus des séances de l'Académie des Sciences, t. 243, p. 1916-1917, séance du 5 décembre 1956 |
La stimulation de certains points de la partie latérale du tegmentum mésencéphalique antérieur, chez le Chat curarisé, produit une réduction d'amplitude de la réponse électrique du noyau cochléaire dorsal à un bref stimulus sonore. Ce mécanisme inhibiteur, contrôlant l'afférence au niveau du premier noyau de relais acoustique, pourrait jouer un rôle dans le déterminisme de l'attention sensorielle.
Des expériences récentes sur le Chat non anesthésié et porteur d'électrodes chroniquement implantées dans le premier relais de la voie acoustique centrale, le noyau cochléaire dorsal, ont mis en évidence des variations remarquables de l'amplitude du message afférent au cours des phénomènes d'attention et d' "habituation" [HernandezPeon, Scherrer et Jouvet (1); Galambos, Sheatz et Vernier (2)]. Nous nous sommes proposé d'explorer en expérimentation aiguë les formations centrales responsables de ce contrôle récurrent.
Les expériences ont été faites sur 21 Chats adultes, immobilisés par un agent curarisant, le Flaxédil ou le Décaméthonium, et soumis à une anesthésie locale à la Novocaïne. L'animal est placé dans un appareil stéréotaxique permettant l'orientation des électrodes de réception et de stimulation. Les réponses sensorielles du nerf auditif (recueillies au niveau de la fenêtre ronde) et du noyau cochléaire dorsal à un bref stimulus sonore ("clic") d'intensité faiblement supraliminaire sont étudiées au moyen d'un oscillographe cathodique à double faisceau. L'activité électrique spontanée du cortex cérébral est par ailleurs enregistrée sur un électroencéphalographe à quatre plumes. Le tronc cérébral est exploré systématiquement, point par point, à l'aide d'une électrode de stimulation du type concentrique bipolaire de fabrication Alvar (distance entre les pôles de stimulation, 0,2 mm; chocs de 1 ms, répétés à 1oo par seconde pendant 1-4 s).
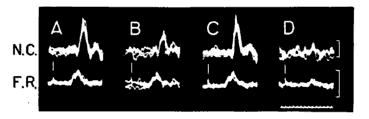
Dans ces conditions, la faradisation de points bien déterminés, dans la partie latérale du tegmentum mésencéphalique antérieur, entraîne une réduction sensible de la réponse du noyau cochléaire au clic, ainsi qu'il est illustré en B. L'effet débute en même temps que la stimulation, et il cesse avec elle (voir C). Le phénomène est stable pour autant qu'on interpose des périodes de repos suffisantes entre les faradisations successives. Plusieurs données expérimentales permettent de penser qu'il s'agit d'un processus d'inhibition vraie et non d'une "occlusion" de la réponse cochléaire au clic par stimulation antidromique de la voie acoustique centrale.
Les expériences étant faites sur l'animal curarisé, et même dans certains cas après enlèvement des osselets de l'oreille moyenne, nos observations ne peuvent s'expliquer par une variation dans la transmission aérienne du stimulus sonore jusqu'à la fenêtre ovale. De plus, il se pourrait que l'effet inhibiteur ne s'exerce pas obligatoirement par la voie acoustique centrifuge reliant l'olive supérieure à la cochlée [Rasmussen (3), Fernandez (4), Galambos (5)]. Comme il apparaît en B, la réponse du nerf auditif (voir F. R.) n'est pas modifiée au moment où celle du noyau cochléaire est nettement réduite. Le phénomène d'inhibition paraît donc s'exercer au niveau du noyau cochléaire lui-même.
Il n'existe pas de relation-simple entre ce contrôle récurrent de l'afférence acoustique et la réaction d'éveil du télencéphale produite par la faradisation de la formation réticulée méso-diencéphalique, et ce pour les deux raisons suivantes : La localisation des points inhibiteurs est beaucoup plus restreinte que celle des régions dont la stimulation provoque l'activation de l'écorce cérébrale. De plus, l'éveil physiologique produit par des excitations sensorielles n'entraîne pas de réduction des messages enregistrés dans les divers noyaux de relais de la voie acoustique, aussi bien chez le Chat " encéphale isolé " [Desmedt et La Grutta (6)] que chez le Chat en expérimentation chronique [Jouvet 7]. On est ainsi conduit à admettre une dissociation entre les processus centraux de la réaction d'éveil, et ces mécanismes inhibiteurs de contrôle qui pourraient entrer en jeu au cours de l'attention sensorielle.
Chat immobilisé, par le Décaméthonium.
Réponses électriques du noyau cochléaire dorsal (N. C.) et du nerf auditif au niveau de la fenêtre ronde (F. R.) à un clic qui est indiqué par une barre verticale. Plusieurs réponses sont superposées de façon à éliminer les fluctuations aléatoires. Étalonnages verticaux : 100 µV. Étalonnage horizontal : millisecondes. A, avant, B, pendant, et C, après la faradisation d'un point inhibiteur au niveau du mésencéphale latéral. D, "masquage" des réponses au clic par un stimulus montrant le caractère neural (et non microphonique) des réponses étudiées.
(1) Science, 123, 1956, p. 331.
(2) Science, 123, 1956, p. 376.
(3) Journal of comparative Neurology, 84 19 46, p. 14
(4) Laryngoscope, 61, 1951, p. 1152.
(5) Journal of Neurophysiology, 19, 1956, p. 49-4.
(6) Journal of Physiology (sous presse).
(7) Journal de Psychologie, 53, 1956, p. 141.