| Le sommeil du 3e type |
| Pierre Magnin Science et Avenir Hors-Série Le Rêve Dec. 96 |
 |
Par Pierre Magnin |
L'approche expérimentale du sommeil est devenue scientifique depuis la mise au point des méthodes d'exploration de l'activité électrique produite par le cortex. Le chercheur a pu quitter alors le domaine de la seule observation personnelle pour entrer dans celui de l'analyse et de la mesure. Le principe de l'électrophysiologie corticale repose sur l'étude des différents potentiels électriques qui existent entre les divers points de la calotte du cortex. Cette technologie a apporté un souffle nouveau à l'analyse du sommeil et du rêve. Aucune méthode expérimentale n'y avait encore donné accès.
Le premier rythme cérébral mais enregistré est le rythme alpha, découvert par le neuropsychiatre allemand Hans Berger en 1924 et publié par lui sous le vocable d'elektrenkephalogram. Ce rythme ne fut confirmé qu'en 1934 - après d'âpres discussions - à Cambridge, à l'occasion d'une démonstration effectuée devant la Société de physiologie de Grande Bretagne. En 1937, le Congrès de Paris consacra son auteur et sa découverte.
Rudimentaire, le premier électroencéphalographe fut vite amélioré. Son champ d'investigation assura l'enregistrement et permit l'analyse de la totalité des points de la calotte crânienne. Lui furent adjoints l'enregistrement des muscles de la face et du menton (électromyogramme ou EMG) et celui des mouvements des globes oculaires (électro-oculogramme ou EOG). L'électroencéphalogramme (EEG) devint polygraphe et se compléta des enregistrements des rythmes respiratoire et cardiaque. L'activité alpha de référence caractérisait un état d'éveil apaisé, le patient gardant les yeux fermés tandis que les autres fonctions restaient normales. Une deuxième activité d'éveil fut identifiée un peu plus tard. Il s'agissait de l'éveil actif et hyperactif pour lequel le sujet gardait les yeux ouverts et exerçait une activité mentale. Ce rythme fut baptisé bêta. L'activité de repos, recueillie pendant le sommeil de nuit, livre un rythme différent de celui de l'état de veille. Il fut appelée delta. Une meilleure maîtrise des techniques et leur affinement par une exploration plus approfondie a permis la mise en œuvre d'une méthodologie séquentielle d'analyse du cortex.
Aujourd'hui, l'amélioration des connaissances et la technologie ont pourvu les polygraphes enregistreurs de divers perfectionnements comme les systèmes vidéo à infrarouge et les magnétoscopes. Le traitement des informations est maintenant assuré par ordinateur. Pourtant, dans les années trente aux États-Unis et à l'aube des moyens techniques, Alfred Loomis et ses élèves furent les premiers à décrypter systématiquement les différentes étapes qui permettent de passer, au cours d'une nuit, de l'état de vigilance aux stades successifs du sommeil. Celui-ci était alors conçu comme un phénomène physiologique global et n'intéressait pas les scientifiques.
De 1934 à 1938, ces chercheurs américains décrivirent cinq stades différents et successifs de déroulement du sommeil : les stades A, B, C, D, et E, qui demeurent les références. L'endormissement succède au rythme alpha de l'éveil calme, yeux fermés. Ce processus traduit une rupture entre l'état de veille et une situation intermédiaire et transitoire qui ouvre vers un enclenchement du sommeil. Il comporte deux étapes A et B qui recouvrent à peu près l'état de somnolence et celui d'assoupissement.
Le stade A est caractérisé par l'émission discontinue de rythmes alpha. Quand s'installe le sommeil, on observe un aplatissement des ondes alpha dont l'amplitude se réduit. C'est le stade B d'assoupissement. Durant cette période, apparaissent chez le sujet les images hypnagogiques. Celles-ci pourraient être des volées hallucinatoires différentes des images entopiques rétiniennes, sans lien avec les voies optiques. Elles parviendraient des centres du névraxe, là où sans doute naissent les images du rêve. Le stade B exprimerait ainsi le "décrochage" du sujet, c'est à dire son isolement sensoriel.
Le stade C commence avec le vrai sommeil. C'est le temps du "sommeil léger" qui est fragile et peut être encore rompu par des stimuli sensoriels. Quant au stade D, il représente le noyau dur de tout le processus du sommeil. C'est autour de ce stade que s'articule le développement physiologique du sommeil. Ce "sommeil profond" est caractérisé par des ondes lentes et par une désynchronisation corticale complète.
Enfin, le stade E, temps du sommeil surprofond, correspond à la période où l'activité métabolique est maximale. Il se caractérise par des trains d'ondes plus lentes.
Ces observations restèrent en l'état bien au-delà de la Seconde Guerre mondiale. Nataniel Kleitman et William Dement reprirent plus tard avec du matériel plus sophistiqué cette classification et en améliorèrent la méthodologie. Ils proposèrent en 1957, une évolution du sommeil en quatre stades notés I, II, III et IV (voir encadrés ci-contre). Parallèlement, Eugen Aserinsky, élève du physiologiste Kleitman, s'appliqua à définir les particularités produites par ces activités insoupçonnées qui accompagnent rythmiquement certains épisodes du sommeil. Il s'agit des mouvements des globes oculaires. On peut les déceler assez systématiquement dès le stade d'endormissement.
En approfondissant son analyse, Aserinsky démontra qu'outre les mouvements des globes oculaires lents et pendulaires caractéristiques de l'endormissement, survient un autre type de mouvements oculaires, rapides cette fois, constitué de phases intenses et brèves. Ils n'apparaissent qu'à certains moments du sommeil léger et de façon surprenante à la fin du sommeil profond, juste avant le réveil. Ils s'accompagnent de trémulations ou de mouvements musculaires cloniques de la face, du menton et des lèvres. Ces mouvements peuvent être complétés par des émissions vocales, mots ou phrases généralement peu compréhensibles. Ce nouveau phénomène fut qualifié de phases de mouvements oculaires (PMO), en français, ou rapid eyes movements (REM), en anglais.
La perspective de lier le phénomène phasique REM ou PMO avec l'état de rêve commençait à poindre dans l'esprit du chercheur et de son équipe. Un autre élève de Kleitman, William Dement, démontra, à partir d'enregistrements pratiqués sur plusieurs centaines de patients, que les deux mécanismes étaient simultanés. En effet, de tous les réveils effectués aux différents stades du sommeil, seuls ceux provoqués au décours d'une phase REM permettaient aux patients de raconter leur rêve. Dement mettait ainsi en évidence la réalité d'un sommeil actif en regard de celui normal ou ordinaire: le sommeil profond, que beaucoup regardaient encore comme un processus purement passif, une sorte de petite mort.
L'annonce de l'existence d'un nouveau type de sommeil, à distinguer du léger et du profond donna le départ à de nombreux travaux qui corroborèrent ceux de Dement. En France, à Lyon, Michel Jouvet entreprit des expériences originales qui montrèrent que chez le chat la privation artificielle de cette phase conduisait à la mort. Jouvet la baptisa, en 1959, sommeil paradoxal.
Paradoxal, il l'est en effet. Ce sommeil à PMO développe une activité cérébrale intense, proche de l'état de l'éveil actif, mais paradoxalement contrebalancée par un relâchement absolu de toute la musculature du corps, de la nuque aux pieds. Ce sommeil, dit du troisième type, fut rapidement interprété comme étant la manifestation électrophysiologique la plus concrète de l'activité corticale survenant au cours du rêve. De plus, toutes les réactions systémiques et neurovégétatives que l'on peut observer à l'occasion d'un rêve étaient actualisées et signifiées pendant le sommeil paradoxal. S'il n'est pas le rêve, le sommeil paradoxal pourrait être considéré comme le meilleur signal des phénomènes qui surviennent au cours du rêve et qui l'accompagnent.
Actuellement, l'investigation des couches neuronales homogènes par recueil de l'activité électrique de surface conduit à rendre possible l'analyse des opérations mentales élémentaires. La détection cohérente de l'espace dans le cadre de l'environnement et du corps devient accessible. C'est le début d'une éventuelle saisie des incidences particulières des modifications projetées au cortex par tous les mécanismes biologiques qui sous-tendent le rêve et qui n'est lui-même qu'un état particulier de l'activité des cellules nerveuses.
Sous observation systématique et contrôlée, un sujet en état de rêve laisse apparaître un état particulier proche du sommeil léger sans y être assimilable. Cet état livre quatre types de signaux fondamentaux : les épisodes REM ou PMO; les effets de sidération musculaire; la production de mots et de phrases peu cohérentes; et chez les humains, le mécanisme d'érection. Tous ces signes peuvent être observés, enregistrés et mesurés à l'occasion d'un réveil provoqué et contrôlé. Leur réalité implique à la fois une activité corticale intense et une participation sensori-motrice et neurovégétative. Le sommeil déconnecte l'organisme de l'influence corticale et l'isole des incidences périphériques et extérieures par le jeu alternatif des groupes de neuromédiateurs; il l'abandonne à la seule loi d'un système nerveux interne et autonome, le neurovégétatif.
Le rêve, lui, semble redistribuer et réajuster le jeu des fonctions organiques et mentales. Le cerveau fonctionne à plein, la pompe cardiaque s'aligne sur cette exigence ainsi que la circulation et la respiration, mais avec une adaptation préférentielle. Ces fonctions réagissent comme lors d'une confrontation événementielle ou émotionnelle.
Cet état nouveau, si différent de celui du sommeil, requiert une participation intense et active des étages sous-jacents au cortex. Voilà pourquoi la phase de sommeil paradoxal s'exprime par un rythme thêta de type hippocampique. La disparition du tonus musculaire, les effets REM ou PMO, les adaptations cardio-vasculaires et respiratoires si caractéristiques de la réaction des centres nerveux signent la participation au rêve des territoires bulbo-ponto-réticulaires et peut-être leur prééminence. Le rêve n'est donc pas l'apanage des seuls humains puisque le cerveau primitif en est partie prenante. On le rencontre à toutes les étapes de la phylogenèse et de l'ontogenèse. Et même si le cheval dort debout, si l'éléphant se balance en dormant, ils sont sujets eux aussi aux phases de sommeil paradoxal.
Les qualités d'acquisition du signal furent les premières et constantes préoccupations de Berger, qui travaillait autant sur le rêve que le sommeil et la pathologie. Elles demeurent aujourd'hui le premier critère de choix de tout centre d'électroencéphalographie, le second critère étant le traitement en temps réel des données recueillies. L'information, la sophistication des systèmes de filtrage parasitaire et d'amplification permettent d'atteindre un premier objectif grâce à ces deux critères. Il reste actuellement un autre objectif à satisfaire : la numérisation avec holter à l'appui des signaux reçus, et dont le traitement des données impose l'usage d'un PC.
Ces progrès technologiques offrent une meilleure approche de l'analyse de l'hypnogramme. Ils permettent de saisir plus finement les différentes phases de sommeil paradoxal dans le déroulement de l'hypnogramme, d'en préciser les entrées, les sorties, les accidents, de les corroborer à d'autres phénomènes. La maîtrise de ces signaux est indispensable pour analyser tous les stades du sommeil, et particulièrement celui du sommeil paradoxal.
Faut-il pour autant penser qu'il existe une correspondance biunivoque entre la vie onirique et le sommeil paradoxal ? On sait que le rêve est articulé à un mécanisme biophysiologique qu'exprime et que manifeste la phase sommeil paradoxal avec ses composantes PMO et neurovégétatives. Toutefois ce stade n'enferme probablement pas l'intégralité du processus dans son contexte d'expression, même si l'on est en droit d'estimer que son support électrique constitue une preuve irréfutable de la réalité d'une physiologie différente de celles de la veille et du sommeil.
Bien que ses mécanismes d'accompagnement soient largement élucidés, la signification du rêve demeure énigmatique.
Les ondes cérébrales
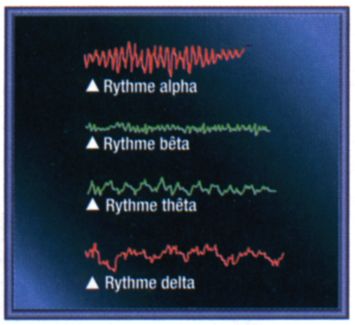
L'électricité cérébrale recueillie à la surface du cuir chevelu se présente sous la forme de rythmes ou ondes cérébrales. On distingue quatre principaux rythmes physiologiques définis par leur fréquence : le rythme alpha (8 à 12 cycles par seconde), qui caractérise l'état de veille calme, (yeux fermés et repos physiologique et mental), le rythme bêta (12 c/s) qui apparaît dans des conditions d'éveil actif et de sommeil, le rythme thêta (4 à 7 c/s) qui apparaît dès l'installation du sommeil et le rythme delta (0,5 à 3 c/s) caractéristique du sommeil lent et profond.
L'analyse électrophysiologique du sommeil
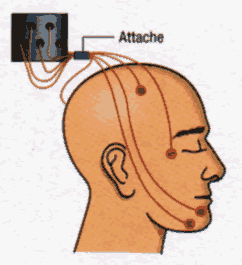
La caractérisation des différents états de veille et de sommeil requiert trois types de signaux électrophysiologiques, enregistrés à l'aide d'électrodes de surface placées à des endroits bien précis: l'électroencephalogramme ou EEG (1), l'électrooculogramme ou EN, (2) pour l'œil gauche et (3) pour l'œil droit et l'électromyogramme ou EMG (4). D'autres paramètres peuvent éventuellement être utilisés pour étayer la classification électrophysiologique, notamment les températures cutanées et rectales, la pléthysmographie pénienne ou le passage de l'air au niveau buccal et nasal.
Le rêveur sous surveillance

Les "laboratoires de sommeil", comme celui du CHU de Besançon, ou ici celui du docteur Rosenberg à l'hôpital Henri-Mondor, à Créteil, comportent en général une ou plusieurs chambres individuelles, confortables et insonorisés. Les appareils d'enregistrement sont situés dans un local sépare afin de ne pas déranger le sujet endormi. Il permet la réalisation de mesures objectives, le contrôle constant des conditions expérimentales et l'évaluation du sommeil de minute en minute grâce aux enregistrements polygraphiques collectés de manière continue tout au long de la nuit.
Stade I: l'endormissement
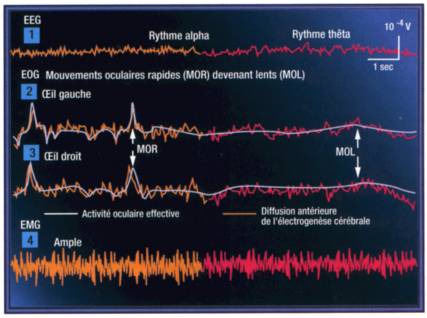
Selon la classification internationale de Kleitman (1), le stade d'entrée dans le sommeil est un stade d'endormissement appelé stade I. C'est une étape importante identifiée par Alfred Loomis en deux moments successifs, A et B. Cet état établit la frontière entre le sommeil et la veille. Il se traduit par un net ralentissement du rythme alpha. Le nombre des cycles par seconde est réduit dans une proportion de 25 à 50 % tandis que croit l'amplitude de chaque fréquence. Ce stade est un mélange de bouffées thêta et bêta. A mesure qu'augmente le besoin de dormir, les mouvements lents des globes oculaires s'accompagnent de la chute des paupières, de la baisse du tonus des muscles de la nuque qui laisse bientôt basculer la tête. Le stade I est une phase courte qui occupe rarement plus de 3 à 5 % de la durée totale du sommeil. La moindre stimulation peut l'annuler et ramener le patient à un état d'éveil immédiat et lucide. Il sert essentiellement à mettre en route les mécanismes biochimiques et physiologiques d'entraînement du sommeil. L'activité de la réticulée mésencéphalique est progressivement inhibée dans son rôle de système activateur ascendant (elle ne stimule le cortex par sécrétion d'acétylcholine qu'après l'avoir été par la noradrénaline que délivre le Locus coeruleus pontique sous-jacent). Ce processus à tiroir doit donc être bloqué pour que le sommeil s'installe et que la réticulée du raphé dorsal sécrète la sérotonine nécessaire.
L'inhibition de sa synthèse crée une insomnie que peut annuler l'injection de son précurseur, le 5-HT. La zone réticulaire du raphé peut être considérée comme le support d'un centre hypnagogique, source des images flashes, sorte de mirage à la frontière du réel et de l'imaginaire.
(1) Manuel de standardisation des états de sommeil chez l'homme, 1938 et 1969
Stade II: le sommeil lent et léger
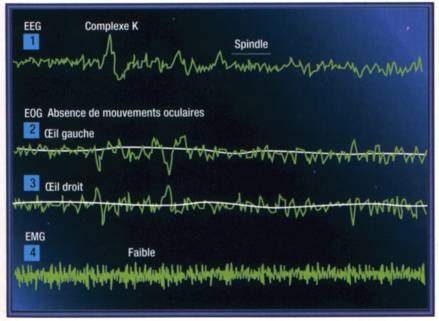
Le stade II est celui d'un sommeil léger mais lent. Il succède à l'endormissement. Avec lui le donneur entre de plain-pied dans un réseau d'aiguillages qui peuvent aussi bien le conduire au réveil qu'au sommeil paradoxal ou au sommeil profond. Le sommeil léger résulte de la mise en œuvre progressive depuis l'endormissement de l'inhibition réticulaire mésencéphalique. Ce temps de sommeil représente à lui seul environ 50 % du total. Son profil EEG est précis: un trace de fond constitué d'une onde irrégulière de 2 à 7 cycles par seconde est interpénétré de fuseaux dont les fréquences sont approximativement le double de celles de l'onde de fond. Ces spindles exprimeraient les mécanismes d'approfondissement du sommeil. S'inscrivent aussi sur le trace de base de grandes ondes lentes, diphasiques, les complexes K qui traduiraient en revanche une réaction aux stimuli extérieurs d'interruption du sommeil. A ce stade, le tonus musculaire n'est pas effacé, le sujet est en hypotonie relative et les grandes fonctions végétatives telles que circulation et respiration sont régulières bien que ralenties, ou plutôt apaisées. Ce stade est celui du réveil facile par effet d'appel, ou de toute stimulation sensorielle.
Les réveils très courts, de moins de trois minutes, sont inconscients mais, fait non négligeable, le mental du dormeur reste impliqué Les résidus d'images hypnagogiques sont toujours des scènes qui se rapportent à la vie éveillée dont le sujet peut se souvenir, surtout s'il se réveille à leur issue. Ce type de sommeil occupe avec l'endormissement environ le premier tiers de la durée du cycle. Ce stade Il est instable: passage obligé de l'éveil à tous les stades du sommeil, il gère de multiples allers-retours consécutifs aux circonstances du sommeil, aux activités physiques, à la santé, à l'humeur, à l'âge et au stress. Le grand vieillard se départit peu de cet état et quitte rarement le stade Il qui représente l'essentiel de sa nuit.
Stades III et IV: le sommeil lent et profond
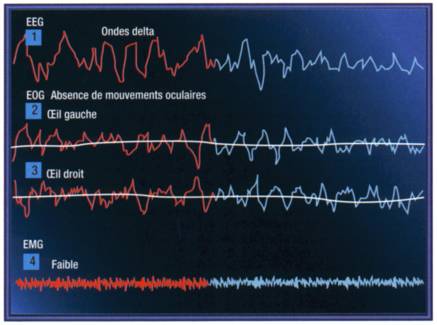
Le sommeil lent, électrophysiologiquement, est le sommeil profond, la référence, longtemps perçu de l'Antiquité à nos jours comme un état totalement passif, une sorte de "petite mort". C'est en réalité une étape importante et très active du sommeil qui répare, régénère et construit. Métabolique et neurovégétatif, ce sommeil efface les fatigues, réhabilite les fonctions et redistribue l'énergie. Il est essentiel chez l'enfant: il recouvre en partie la période de sécrétion de l'hormone de croissance, qui active dès le petit matin tous les processus de synthèse, en particulier protéiques. Ce type de sommeil fait aussi le lit du rêve en préparant l'émergence du sommeil paradoxal à chacune des ruptures périodiques du cycle. Phénomène remarquable, surtout chez l'adulte qui rêve moins que l'enfant, le sommeil lent paradoxal est presque totalement accompli dans les premières heures de la nuit en recouvrement des trois ou quatre premiers cycles. Son effacement progressif dans la seconde moitié de l'espace de sommeil explique l'apparition et la facilitation des insomnies de l'adulte et surtout des personnes âgées. Son enregistrement inscrit une onde très lente d'activité delta, de l'ordre de deux cycles par seconde. De grande amplitude, elle est tout d'abord inégale et discontinue (stade III) avant de se régulariser progressivement (stade IV). On observe à ce niveau une accentuation du degré d'hypotonie du donneur, c'est-à-dire une évolution vers l'éveil. Les phases du sommeil lent paradoxal coïncident aussi avec une activité métabolique de production neuronale d'amines spécifiques du sommeil. L'expérimentation de ces peptides, en particulier du DSIP (Delta Sleep Including Peptide), a montré qu'ils induisaient plutôt une activité de régulation des rythmes de sommeil qu'un effet hypnotique. Le sommeil lent paradoxal représente le centre actif de l'hypnogramme. Il est avec le sommeil paradoxal l'expression des fonctions régénératrices de sommeil et du rêve.
Stade V: le sommeil paradoxal
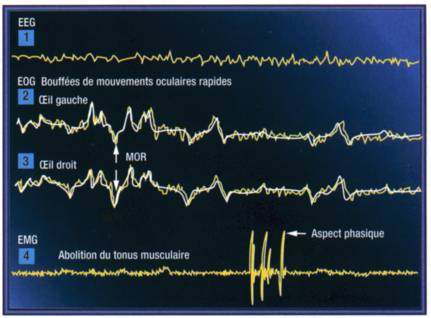
Au stade V, à l'issue du sommeil lent paradoxal (encore appelé stade PMO, phase de mouvements oculaires, ou REM, rapid eyes movements), L'EEG affiche soudain une activité plus rapide traçant "une onde en dents de scie" de très bas voltage. Ce stade ressemble, électriquement parlant, au stade 1. En parallèle, l'EMG traduit une abolition de l'activité musculaire tandis que les mouvements oculaires s'inscrivent en salves phasiques nombreuses. Rythme cardiaque et respiration deviennent irréguliers signant ainsi la dissociation neurovégétative. L'érection s'installe, et divers accès cloniques de la face, des pieds et des doigts peuvent survenir. Comparable au stade 1, le tracé du stade V le situe proche de l'état de vieille alors que cette phase de sommeil profond original et particulier requiert de fortes stimulations afin d'accepter le réveil du donneur. Le stade V représente entre 20 et 25 % de la durée totale du cycle, selon l'âge et l'état du sujet. Il s'inscrit en épisodes brefs après chaque développement d'un cycle des stades de sommeil. Ses épisodes sont de plus en plus longs et de plus en plus fréquents à mesure que la durée du sommeil s'épuise et que la nuit s'avance. Son délai d'apparition est de l'ordre de 90 minutes après l'endormissement. A la différence des "états intermédiaires", qui encadrent tous les autres stades de sommeil et pour lesquels il n'existe pas de frontières nettes, la survenue du sommeil paradoxal tranche avec celle du sommeil lent profond qui le précède. Un véritable coup de feu éclate dans le silence du sommeil lent profond. Les mouvements oculaires apparaissent tandis que restant en mydriase, la pupille signe la persistance du sommeil. Ces mouvements sont déclenchés par le lancement de l'activité ponto-genouillée-occipitale. Ils pourraient accompagner les images du rêve qui peuvent résulter d'une stimulation du cortex occipital. Le cerveau est actif mais désynchronisé du reste du corps. Ces épisodes témoignent de l'activité intense et de l'importance du rêve pour la qualité de la vie psychique, intellectuelle et biologique de l'homme.