| Etude des mécanismes du sommeil physiologique |
| M. Jouvet, J. Dechaume et F. Michel Lyon Médical N 38 -18 Septembre 1960 |
Introduction
Les progrès de la neurophysiologie ont vu la confrontation de deux hypothèses principales concernant le sommeil:
- a) ou bien le sommeil est dû au relâchement passif d'un système de vigilance;
- b) ou bien il est dû à l'intervention active d'un système hypnique.
Il semble que ce soit à Gayet, médecin de l'Hôtel-Dieu de Lyon, en 1875, que l'on doit la première description d'un cas d'encéphalite lésant le troisième ventricule, accompagnée d'une somnolence invincible et de paralysies oculaires. Cette constatation laissait entrevoir l'existence de structures nerveuses responsables de l'état de veille.
Au cours de la grande épidémie d'encéphalite de 1918, Von Economo situa également, à la partie postérieure du troisième ventricule, et au niveau de la partie haute du tronc cérébral, les lésions responsables de l'encéphalite léthargique. C'est donc au niveau de l'hypothalamus que les physiologistes établirent des lésions expérimentales en traînant de la somnolence (Ranson et son école, 1934). Raphaël Dubois (1894) semble avoir été un précurseur en localisant, chez la marmotte, au niveau du plancher de l'aqueduc, le siège des processus du réveil. Il devenait alors évident que certaines parties du tronc cérébral étaient responsables de l'état de vigilance et c'est le mérite de Moruzzi et Magoun et de leur école, reprenant les expériences classiques de Bremer (1936) d'avoir montré que la formation réticulée du tronc cérébral constituait un système activateur ascendant responsable de l'état de veille.
Cependant, les partisans de l'existence d'un système de veille se montrent extrêmement discrets lorsqu'il s'agit d'expliquer son ´relâchementª passif. Ils invoquent alors une fatigue neuronique ou une déafférentation: un mécanisme hypothétique empêchant l'arrivée des signaux du monde extérieur au sein de la formation réticulaire. Cette ´déafférentationª avait déjà été proposée par Lépine (1895) qui émit l'idée que le sommeil naturel pouvait être causé par le retrait des prolongements de certaines cellules nerveuses. Cependant, aucune expérience n'a permis de prouver, électrophysiologiquement, l'existence d'une déafférentation réticulaire au cours du sommeil physiologique.
C'est donc à quelque mécanisme inhibiteur qu'il appartient de freiner, de façon active, le système réticulaire activateur au cours du sommeil. Mais la délimitation de systèmes ou de centres hypnogènes actifs est plus difficile à réaliser que celle d'un système d'éveil, car nous comprenons encore mal les mécanismes d'inhibition nerveuse.
Pavlov fut le premier à insister sur l'importance primordiale du cortex comme facteur d'inhibition des centres nerveux sous-jacents Il montra qu'au cours de certains conditionnements complexes qui mettaient sûrement les cellules corticales en jeu, le sommeil apparaissait de facon active sinon explosive. Plus tard, Hess, en stimulant le thalamus, put induire le sommeil chez le chat, et apporta ainsi des arguments en faveur de l'existence de mécanismes inhibiteurs.
L'électroencéphalogramme (E. E. G.) est venu enrichir, considérablement les techniques d'étude du sommeil:
Dès les premières études, il est apparu que les aspects E.E.G. du sommeil étaient très complexes. La majorité des auteurs s'accorde cependant pour décrire quatre stades principaux chez l'homme: Blake, Gérard et Kleitman (1939), Davis (1938), Loomis (1938), Dement et Kleitman (1957). Des travaux récents ont attiré l'attention: Aserinsky et Kleitman (1955), Dement et Kleitman (1957), sur un stade particulier du sommeil accompagné de mouvements rapides des yeux et en relation probable avec le rêve.
Chez le chat: après les études classiques de Rheinberger et Jasper (1937) et de Hess (1953) qui ont révélé que le sommeil physiologique s'accompagnait de fuseaux et d'ondes lentes corticales et sous-corticales, l'existence de périodes d'activité corticale rapide pendant le sommeil fut ensuite reconnue (Rimbaud, Passouant et Cadilhac, 1955), tandis que Dement (1958) montrait que cette activité rapide s'accompagnait de mouvements des yeux et pouvait ainsi traduire une activité onirique; pour cet auteur, le stade rapide du sommeil chez le chat représenterait une phase de sommeil activée (activated sleep), intermédiaire entre le sommeil profond, caractérisé par les ondes lentes, et l'état de veille.
1 - Rappel méthodologique
L'expérimentation animale, associant destruction et stimulation nerveuse, permet seule de résoudre le problème des structures nerveuses responsables des différents stades du sommeil. On sait que deux voies d'étude ont été ainsi suivies:
La première (Bremer, 1936; Batini, Moruzzi, Palestini Rossi, Zanchetti, 1958) se base essentiellement sur le tracé E.E.G. cortical qu'elle considère comme le témoin le plus fidèle du niveau de vigilance. Elle arrive ainsi à assimiler tracé lent et sommeil, tracé rapide et éveil. Il existe, cependant, de nombreuses exceptions:
- Un tracé "synchronisé" ou lent n'est pas forcément un signe de sommeil physiologique. Tels la narcose au penthiobarbItal. ou le coma par destruction de la formation réticulée (French et Magoun, 1952), dont l'irréversibilité permet d'éliminer un phénomène de sommeil.
- Un tracé rapide n'est pas forcément signe d'éveil: la narcose à l'éther, par exemple, s'accompagne d'une "activation" corticale et il existe des phases de sommeil avec activité corticale rapide chez le chat.
- Enfin et surtout, si l'on étudie, en expérimentation chronique, un animal sectionné entre les tubercules quadrijumeaux, la seule considération de son tracé cortical ferait affirmer qu'il dort alors que l'animal peut se tenir sur ses pattes et réagir à la douleur par des mouvements de fuite et des miaulements.
La nécessité impérieuse d'une vérification du comportement de sommeil de l'animal nous a conduit à enregistrer l'activité électrique corticale et sous-corticale, en corrélation avec les manifestations somato-végétatives du sommeil (Lhermitte et Tournay, 1927; Tournay, 1910; Piéron, 1913), (en particulier l'activité tonique musculaire, les rythmes cardiaques et respiratoires, etc...). Dans le cas de section du tronc cérébral, l'activité électrique de la partie de l'encéphale située en arrière de la lésion (et non seulement en avant), donc en liaison avec le milieu extérieur par ses afférences et efférences principales, était enregistrée au cours des variations comportementales périodiques de vigilance (fig. 1). Il est, en effet connu depuis Goltz (1892), qu'un animal décortiqué peut dormir. Les travaux de Bard (1958) ont également montré que l'animal mésencéphalique présentait des alternances comportementales de veille et de sommeil.
2 - Aspects E.E.G. du sommeil physiologique chez le chat
Nous résumerons ici des résultats expérimentaux (Jouvet et Michel, 1958, 1959, 1960; Jouvet, Michel et Courjon, 1959) qui nous conduisent à proposer l'existence de deux systèmes: télencéphalique et rhombencéphalique, responsables des stades d'ondes lentes et du stade d'activité rapide au cours du sommeil. L'activité électrique cérébrale recueillie par des électrodes corticales ou sous-corticales implantées à demeure chez plus de cinquante chats chroniques (c'est-à dire étudiés plus de cinq jours, au cours du sommeil physiologique) fut mise en corrélation avec l'activité électromyographique des muscles de la nuque, les mouvements des yeux, le rythme cardiaque et la respiration. Ces investigations furent pratiquées soit chez des chats intacts, soit chez des chats décortiqués totaux ou subtotaux, soit chez des animaux porteurs de sections du tronc cérébral à des niveaux variables: animaux mésencéphaliques ou pontiques chroniques. Enfin, des expériences de contrôle furent réalisées chez des animaux chez qui le cervelet fut enlevé totalement.
A. - Chez le chat endormi
On peut distinguer deux stades électriques bien différents:
1 - Endormissement et sommeil "lent": ce stade est marqué par l'apparition de fuseaux qui envahissent le cortex, le diencéphale et la formation réticulée mésencéphalique (F. R.). Ensuite le sommeil lent est caractérisé par l'apparition, au niveau du cortex et des structures diencéphalo mésencéphaliques, d'ondes lentes de deux à quatre c/sec., de haut voltage, alors que l'activité hippocampique ventrale présente des pointes très brèves. L'apparition des ondes lentes mésencéphaliques se fait toujours après un certain retard avec l'apparition des ondes lentes corticales.
Au cours de ce stade, l'animal se met en posture de sommeil; les membranes nictitantes viennent recouvrir se pupilles qui sont en myosis. La nuque fléchit jusqu'à ce que repose la tête, tandis que l'activité électrique des muscles de la nuque diminue discrètement. La respiration est régulière ainsi que le rythme cardiaque qui se ralentit par rapport à l'état de veille. Au fur et à mesure de l'invasion de la formation réticulaire par des ondes lentes, le seuil d'éveil par stimulation directe de la formation réticulaire augmente de façon significative (20 à 50 %)
2 - "La phase paradoxale" (P. P.): Elle fait toujours suite à une phase de sommeil lent et n'apparaît jamais d'emblée après un tracé de veille. Elle débute soudainement et se caractérise par une activité cortico-mésodiencéphalique rapide de bas voltage, identique à celle de l'éveil, alors que les formations rhinencéphaliques dorsales peuvent parfois présenter une activité rythmique lente, identique à celle qui fut décrite au cours de l'éveil à ce niveau. En même temps, apparait, au niveau de la F. R. pontique, une activité de fuseaux de six à huit c./seconde, de haute amplitude, qui apparaît avec l'activité rapide et disparaît avec elle. Cette phase, d'une durée de dix à quinze minutes, s'accompagne immédiatement d'une disparition totale de l'activité E.M.G. des muscles enregistrés. Soit spontanément, soit sous l'influence d'une stimulation extérieure, l'activité électrique corticale peut revenir au stade antérieur du sommeil avec réapparition de fuseaux ou d'ondes lentes, ou bien il peut y avoir réveil de l'animal.
Au cours de cette P. P., la posture de l'animal est celle du sommeil; il n'y a plus aucun tonus musculaire postural et si la tête ou un membre de l'animal sont en surplomb, ils pendent alors, flasques; les membranes nictitantes recouvrent presque entièrement les pupilles qui sont en myosis, tandis que les globes oculaires sont fréquemment animés de brèves secousses rapides de type nystagmiforme, surtout dans le sens latéral.
Il existe, également, des mouvements des vibrisses, plus rarement de brèves secousses des mâchoires et de la queue. Des variations cardio-respiratoires apparaissent constamment au cours de ce stade: le rythme respiratoire devient irrégulier, plus superficiel et plus rapide qu'au cours de la phase d'ondes lentes, tandis que le rythme cardiaque, selon les animaux et selon les P. P. peut subir une accélération nette ou plus souvent, au contraire, se ralentir.
Le seuil d'éveil, en décibels, par une stimulation auditive augmente au cours de ce stade par rapport au stade d'ondes lentes. Il en est de même pour le seuil d'éveil par stimulation directe de la formation réticulée mésencéphalique (élévation de 200 à 300 C). Enfin, on remarque une diminution nette des réponses évoquées corticales et surtout réticulaires, soit par rapport au stade de fuseaux du sommeil, soit par rapport au stade d'éveil accompagné d'une activité rapide identique.
Cette phase, que nous avons retrouvée chez tous les animaux, se répète périodiquement au cours du sommeil comportemental, séparée par des intervalles de vingt à trente minutes (voir fig. 15).
B. - Chez le chat néodécortiqué chronique
(fig. 4):
Cette mutilation provoque un changement net et sur prenant de l'E.E.G. recueilli au niveau des structures sous corticales. Il existe, en effet, une absence permanente de fuseaux ou d'ondes lentes au cours de la survie de l'animal (jusqu'à trois mois). L'activité des formations centrales mésodiencéphaliques présente, de façon continue, une activité rapide et de très bas voltage, quel que soit l'état de vigilance de l'animal. Seul l'enregistrement au niveau de l'hippocampe permet alors de retrouver un stade électrique correspondant au sommeil profond: pointes brèves de haut voltage qui font suite à la période rapide de l'éveil. Par contre, la P. P. se manifeste par les mêmes phénomènes que chez l'animal intact: apparition de fuseaux à 6 c./seconde de haut voltage au niveau de la F. R. pontique, activité rapide rhinencéphalique, tandis que l'activité E.M.G. disparaît et qu'apparaissent des modifications cardio-respiratoires et des mouvements des yeux.
C. - Chez l'aninal chez qui il reste une surface néocortex minime
(le cortex frontal en particulier): on retrouve les deux phases E.E.G. décrites chez l'animal normal.
D. - De même chez l'aninal cérébellectomisé
présente les mêmes caractéristiques E.E.G. et comporte mentaux au cours du sommeil que l'animal normal, et il est surprenant de constater au cours de la P. P. une disparition totale de la rigidité de type alpha qui fait suite à une telle opération.
E. - Aninal mésencéphalique chronique
L'activité électrique des formations corticales et diencéphaliques situées en avant de la section présente continuelle ment l'aspect classique décrit au niveau du cerveau isolé (Bremer, 1935), c'est-à-dire un mélange de fuseaux et d'ondes lentes continues, encore que de brèves phases d'activation apparaissent au niveau du cortex lors de stimulations olfactives.
En arrière de la section par contre, l'activité mésencéphalique reste constamment rapide et de bas voltage et les ondes lentes n'y apparaissent plus; seule peut s'objectiver la P. P.: apparition périodique de fuseaux au niveau de la E. R. pontique et disparition totale de l'activité E.M.G. de la nuque, d'autant plus remarquable que l'activité musculaire est augmentée lors de la veille (hypertonie de décérébration). Chez les préparations sectionnées à la limite antérieure du mésencéphale, les mouvements des yeux sont moins nets que chez les préparations décortiqués ou normales. Par contre, après une section à la partie moyenne du mésencéphale, il n'existe plus de mouvements des yeux au cours de la P. P.
Cependant chez toutes ces préparations, on remarque d'importantes variations cardio-respiratoires: accélération de la respiration et ralentissement du rythme cardiaque au cours de la phase paradoxale. Enfin, la durée de ces phases est analogue à celle présentée par l'animal intact: dix à quinze minutes; leur intervalle est, par contre, plus long (quarante à soixante minutes).
F. - Aninal pontique
Chez une préparation sectionnée selon un plan incliné à la partie antérieure du pont (préparation rostro-pontique), les phénomènes identiques à ceux de l'animal mésencéphalique s'observent. Par contre, chez les chats sectionnés au milieu du pont et en avant de l'arrivée du trijumeau (medio pontine pretrigeminal preparation, M. P. P.) (Batini et collaborateurs, 1958), deux phénomènes très particuliers apparaissent: en arrière de la section, il n'existe plus aucune variation périodique de l'activité musculaire, cardiaque ou respiratoire, analogue à celle présentée par l'animal mésencéphalique. Une activité E.M.G. persiste constamment au cours de la survie de l'animal (jusqu'à dix jours). En avant de la section par contre, l'activité corticale est très fréquemment rapide, comme l'ont montré les observations de l'école de Moruzzi (1958). L étude attentive des mouvements des yeux (sous la dépendance du III et du IV, situés en avant de la section) montre qu'il existe des stades d'activité corticale rapide au cours desquels l'animal peut suivre de façon verticale un objet mouvant, tandis qu'il existe d'autres stades d'activité rapide au cours desquels il ne suit plus un objet mais où ses yeux sont animés de mouvements spontanés, verticaux également, et que nous interprétons comme traduisant une activité rapide de sommeil. Il nous a été possible, en effet, au cours de ces périodes, de "réveiller" la partie rostrale de l'encéphale par une stimulation olfactive et de voir réapparaître des mouvements de fixation des globes oculaires, sans que l'activité électrique cérébrale ait changé.
3 - Topographie des systèmes responsables des deux stades du sommeil
A. - Première phase de sommeil (fuseaux et ondes lentes)
Le système neuronique responsable de cette phase nécessite obligatoirement la présence du néocortex puisqu'il n'y a ni fuseaux, ni ondes lentes sous-corticales chez l'animal décortiqué, ou en arrière d'une section du tronc cérébral.
L'étude d'animaux porteurs de sections partielles du tronc permet de préciser que les voies descendantes inhibitrices corticifuges sont diffuses au sein de la formation réticulaire du tronc cérébral car des ondes lentes apparaissent en arrière d'une section, laissant une partie, même minime, de la formation réticulaire intacte.
B. - Phase paradoxale
Le mécanisme responsable du déclenchement de la P. P. se situe obligatoirement à la partie postérieure du mésencéphale et à la partie antérieure du pont (N. pontis oralis), car des animaux sectionnés en avant de ces structures continuent à présenter les manifestations périphériques de la P. P. tandis que la préparation sectionnée au milieu du pont (M.P.P.) ne présente plus de telles variations (fig. 7).
Par contre, les mécanismes responsables de l'activité rapide corticale au cours de cette phase semblent différents de ceux de l'éveil. Il est apparu, en effet, que des animaux dont la F.R. est totalement détruite au niveau du tegmentum mésencéphalique (fig. 8) et qui sont incapables d'un éveil cortical par stimulation nociceptive, continuaient à présenter, cependant, une ´activation corticale au cours de la P. P. si la partie ventrale du pied du mésencéphale est respectée (au niveau du noyau interpédonculaire et des corps mamillaires) (fig. 9). Or, on sait que de telles structures sont des voies de passage d'un circuit (limbic mid brain circuit, Nauta, 1958; Nauta et Kuypers, 1958) unissant certaines formations situées à la partie rostrale du pont et à la partie caudale du mésencéphale (noyau de Betcherew et de Gudden) aux formations limbiques et rhineneéphaliques. Il est fort probable qu'un tel circuit soit responsable des aspects électriques corticaux de la P.P.
Les phénomènes somato-végétatifs qui accompagnent la P.P. doivent s'expliquer par la mise en jeu active de la F. R. inhibitrice ponto-bulbaire (Magoun, 1950) dont on connaît le rôle de contrôle généralisé sur le tonus musculaire. A ce niveau, se situent également des centres respiratoires et cardio-vasculaires dont la mise en jeu explique les variations caractéristiques des rythmes respiratoires et cardiaques.
4 - Les expériences de stimulation
Les expériences de stimulation confirment la dualité des mécanismes responsables du sommeil (fig.10 et 11).
1 - On sait, depuis Hess (1928), que la stimulation à basse fréquence des parties médianes du thalamus peut provoquer une synchronisation de l'E. E. G. chez le chat, et il est probable qu'une telle stimulation mette en jeu le système diffus cortical inhibiteur qui vient agir sur le système réticulaire ascendant activateur; nous avons, nous-mêmes, pu reproduire fréquemment une telle synchronisation et un comportement d'endormissement par stimulation de la massa intermedia du thalamus à basse fréquence. Par contre, la stimulation du thalamus ne nous a jamais permis de provoquer de phase paradoxale du sommeil.
2 - Il en est tout autre lors de la stimulation des parties basses du tronc cérébral (partie postérieure de la F. R. mésencéphalique, parties antérieure et moyenne de la formation réticulée pontique). C'est ainsi que chez des animaux intacts, il a été possible de provoquer l'apparition de phase paradoxale typique avec tous ses caractères E.E.G. et périphériques, en stimulant certains points des formations mésencéphaliques ou pontiques, à condition que cette stimulation ait lieu au cours d'une période d'ondes lentes (fig. 10)
Chez des animaux mésencéphaliques (fig. 11), la stimulation d'endroits limités des F. R. mésencéphaliques postérieurs et pontiques est également capable de déclencher des périodes durables d'archéo-sommeil avec toutes ses caractéristiques: activité rythmique pontique, disparition de l'activité E.M.G., mouvements rapides des yeux, variations respiratoire et cardiaque, etc...
Des phases d'une quinzaine de minutes ont ainsi pu être obtenues après des stimulations d'une durée de une seconde. Après réveil spontané de l'animal, la même stimulation entraîne, au contraire, une phase d'hypertonie avec agitation; aussi, n'a-t-il jamais été possible de déclencher à nouveau une période ´ d'archéo-sommeil ª immédiatement après réveil; il fallait toujours attendre quinze à trente minutes pour qu'une nouvelle excitation provoque alors une phase de sommeil. Chez d'autres animaux, ces mêmes phénomènes ont été obtenus par la sommation d'excitation de longue durée et, dans certains cas, une latence de une à deux minutes était observée entre la fin de la stimulation et l'apparition de l'archéo-sommeil.
Des contrôles ont montré que ces phénomènes ne s'accompagnaient pas de variations significatives de la tension artérielle. L'atropine, à la dose de 0,2-0,3 milligramme par kilogramme diminue considérablement la durée des P. P. ou même empêche leur apparition, aussi bien chez l'animal intact que chez l'animal mésencéphalique chronique. Par contre, l'injection de drogues cholinergiques (esérine) a permis de montrer que l'on pouvait obtenir des phases paradoxales significativement plus longues, mais dont la fréquence n'était pas augmentée (fig. 12).
5 - L'ensemble de ces résultats
L'ensemble de ces résultats conduit donc à admettre l'intervention de deux systèmes différents au cours du sommeil physiologique (fig. 13):
1 - Le premier système entrerait en jeu au cours de l'endormissement (fuseaux) et au cours du sommeil (ondes lentes) de l'animal intact. Ce système nécessite le néocortex responsable de l'activité lente. Cette activité lente corticifuge traduit un phénomène d'inhibition car le seuil d'éveil par stimulation directe de la formation réticulaire s'élève lors de ce stade. On peut donc parler légitimement de sommeil télencéphalique à propos de la phase de fuseaux et d'ondes lentes du sommeil. La mise en jeu physiologique d'un tel sommeil peut être provoquée par la répétition de stimuli, soit indifférents, soit forts, comme dans le cas de l`inhibition supra-liminale pavlovienne. Il est possible que ce stade télencéphalique représente un stade acquis au cours de la télencéphalisation et qu'il traduise l'expression d'un néo-sommeil.
Sur cette seule phase de sommeil, il a pu être mis en évidence des phénomènes plastiques d'apprentissage négatif du type d'habituation de la réaction d'éveil (Jouvet, 1959, 1960).
2 - Le stade rapide du sommeil, que nous avons appelé phase paradoxale dépend d'un système totalement différent:
Il existe, en effet, à la partie caudale du tronc cérébral (F. R. mésencéphalique postérieure, F. R. pontique antérieure, noyau reticularis pontis oralis, noyau de Betcherew) un système neuronique dont la mise en jeu est responsable de ce stade rapide du sommeil chez le chat. Ce système tient sous sa dépendance, par l'intermédiaire de la formation réticulée inhibitrice, les phénomènes somato-végétatifs caractéristiques de cette phase (disparition de toute activité tonique musculaire même dans des cas de rigidité de décérébration ou de décérébellation, variations des rythmes respiratoires et cardiaques). Les mouvements des yeux, qui caractérisent ce stade, sont sous la dépendance de la F. R. mésencéphalique antérieure. Ce système entre en rapport, en avant, avec des fibres à projection rhinencéphalique qui sont responsables de l'activité rapide corticale, car il semble que celle-ci ne soit pas sous la dépendance directe du système réticulé activateur ascendant. Le fait qu'il ait été possible de mettre en jeu, dans certaines conditions, la phase paradoxale du sommeil par stimulation directe ne préjuge en rien de son déclenchement physiologique qui reste encore peu connu à l'heure actuelle. Cependant, l'existence d'une phase réfractaire de dix à quinze minutes chez l'animal intact, de vingt-cinq à trente minutes chez l'animal mésencéphalique chronique, après l'apparition de chaque phase paradoxale, soulève le problème d'un mécanisme neuro-humoral. Enfin, l'action des drogues, antagoniste de l'atropine et favorisante de drogues cholinergiques, laissent supposer que ce mécanisme neuro-humoral serait cholinergique. Ce stade de sommeil, qui existe seul chez des chats chroniques mésencéphaliques ne possédant que la portion archaïque de l'encéphale, pourrait ainsi être assimilé à un archéo-sommeil.
6 - Aspects E.E.G. du sommeil physiologique chez l'homme
A.- Sujets normaux
Nous avons pratiqué quinze tracés de sommeil (de 22 heures à 6 heures du matin) sur quatre sujets adultes: deux hommes et deux femmes de 25 à 35 ans. De façon continue, l' E. E. G. était enregistré au niveau du scalp ainsi que les mouvements des yeux, grâce à des électrodes placées de part et d'autre des globes oculaires, l'activité musculaire au niveau de la nuque ou des sterno-cleïdo mastoïdiens, le réflexe psycho-galvanique, l'électrocardiogramme et enfin la respiration grâce à un système placé autour du thorax.
Dans certains cas, le seuil d'éveil fut testé par stimulations auditives pratiquées par l'intermédiaire d'un haut parleur déclenché par stimulateurs à voltages variables. Lors des éveils provoqués au cours des différents stades de sommeil, il était demandé au sujet s'il se souvenait d'un rêve.
Les sujets étaient couchés confortablement dans un lit situé dans une pièce faiblement éclairée, l'observateur restant devant l'électroencéphalogramme, séparé du sujet par l'intermédiaire de plusieurs rideaux ou dans une pièce contiguë. A tout instant, il était possible d'observer le sujet sans faire de bruit.
Il est vite apparu, au cours de ces enregistrements, que l'index électromyographique était mauvais chez l'homme, car aussi bien l' E. M. G. des muscles de la nuque que celui des sterno-cleïdo-mastoïdiens disparait dès les premières phases de l'endormissement, du fait de la posture de repos et de sommeil chez l'homme.
L'enregistrement de l'orbiculaire des paupières s'est révélé également difficile chez le sujet normal. Il en est de même avec notre technique d'enregistrement du réflexe psycho-galvanique, dont nous ne ferons pas état.
Résultats
Nous adopterons la terminologie de Dement et Kleitman (1957) au sujet des différents stades du sommeil, pour n'insister que sur la phase paradoxale (fig. 14a).
Stade 1: bas voltage avec une activité relativement rapide et un manque absolu de fuseaux. Il s'observe immédiatement après l'endormissement et peut être extrêmement court chez certains sujets; il est totalement dénué de mouvements oculaires et le seuil d'éveil est extrêmement bas.
Stade 2: présence d'une activité de fuseaux apparaissant sur un arrière fond d'activité rapide et de bas voltage. Par fois, il peut déjà exister une activité thêta. C'est à ce stade que sont recueillis les K complexes au niveau des régions antérieures.
Stade 3: intermédiaire, caractérisé par l'apparition d'ondes lentes de haut voltage auxquelles s'associent encore quelques fuseaux.
Enfin, le stade 4 est caractérisé par la présence d'ondes lentes delta de grande amplitude.
La phase paradoxale chez l'homme (fig. 14b)
Elle se caractérise:
a) Au point de vue E.E.G., par une activité de bas voltage, une absence totale de fuseaux et une absence totale de K complexes lors des stimulations auditives. Au niveau des régions postérieures, il existe parfois une activité identique à celle de l'alpha avec cependant une amplitude moins grande. Parfois, à cette activité, s'associe une activité plus irrégulière, mais toujours de bas voltage à 5-6 c./seconde. On ne remarque jamais une activité très rapide, comme on peut la voir durant la réaction d'arrêt. Au niveau des régions antérieures, par contre (dérivations fronto-temporales ou vertex-pariétales ou fronto-vertex), on remarque fréquemment la présence d'ondes lentes à 3 ou 4 c./seconde ayant une allure triangulaire ou quadrangulaire, ´ en dents de scie ª. Ces ondes lentes ne sont pas contemporaines des mouvements des yeux car elles ne se retrouvent pas dans les dérivations frontales où apparaissent alors les artefacts oculaires. Ces phases paradoxales, d'une durée de trente à quarante minutes, se sont répétées de deux à quatre fois dans une nuit, selon les sujets (fig. 15). Les intervalles minima qui les séparaient ont toujours été supérieures à trente minutes. Parfois, il est arrivé que les sujets réveillés au cours d'un stade paradoxal se replongent, après rendormissement. très rapidement dans ce même stade.
b) Phénomènes périphériques (fig. 16): les mouvements des yeux sont très caractéristiques et ressemblent, en tous points, à ceux qui sont observés chez le chat et qui ont été décrits par Dement et Kleitman chez l'homme. Mouvements rapides, de type nystagmiforme, irréguliers, apparaissant par bouffées, ils marquent l'apparition de la phase paradoxale et en signalent la fin par leur disparition. A ce stade. le tonus musculaire de l'orbiculaire disparaît car les paupières s'entrouvent pour laisser apparaître la sclérotique blanche. Il est fréquent, également, de constater des mouvements discrets des commissures labiales et des doigts.
Les variations respiratoires sont constantes chez tous les sujets: irrégularité, diminution de l'amplitude et accélération qui contrastent avec la régularité observée au cours des autres stades du sommeil. Les variations du rythme cardiaque sont moins constantes: l'irrégularité, en général, est manifeste, et, dans 60 % des cas, nous avons observé une accélération du pouls, et dans 20 %, un ralentissement; chez un sujet cependant, il n'y eut pas de modification, sauf lors des dernières P. P. au cours de la nuit.
Cinquante réveils ont été provoqués au cours des tracés de sommeil: sur vingt réveils provoqués au cours de la P.P., douze fois les sujets déclarèrent qu'ils avaient rêvé. Sur trente-cinq réveils provoqués en dehors de la phase paradoxale, au cours des stades 1, 2, 3 et 4, un seul sujet déclara une fois avoir rêvé.
B.- Le sommeil au cours d'un syndrôme de décortication chronique
L'analyse structurale de l' E.E.G. humain souffre d'un handicap majeur: l'impossibilité de réaliser des lésions expérimentales. Cependant, dans certains cas, la maladie peut réaliser des atteintes nerveuses systématisées comparables à celles de l'expérimentation. C'est le cas d'une malade de 30 ans, hospItal.isée à la Clinique Neuropsychiatrique, présentant, depuis trois ans et demi, un tableau clinique de décortication (syndrome d'aperceptivité réactive hyperpathique et hypertonique), (Dechaume et Jouvet, 1960), consistant en une absence totale de perceptivité visuelle ou auditive, avec conservation de réactions végétatives et mimiques à la stimulation douloureuse et présentant une attitude posturale en flexion des membres supérieurs avec un réflexe de Magnus et Klein.
Chez cette malade, la ventriculographie objective une dilatation progressive et globale des ventricules (fig. 17). Des tracés ont permis de retrouver, chez cette jeune femme, un rythme de sommeil analogue à celui du chat décortiqué (fig. 18). L'état de veille se caractérise par une très nette hypertonie des fléchisseurs du membre supérieur, se traduisant par une activité E.M.G. intense recueillie au niveau du biceps, par des mouvements incessants de mâchonnement. Fait caractéristique, il n'apparaît jamais aucune onde lente de haut voltage ou fuseau, ni K complexe sur l' E.E.G.: comme chez le chat décortiqué chronique, le tracé E.E.G: reste rapide et de bas voltage de façon permanente quel que soit l'état de vigilance (fig. 19). Le stade paradoxal est typique: l'activité E.M.G. disparaît alors totalement, tandis que les mouvements des yeux sont identiques à ceux que présente un individu normal au cours de ce stade. Il existe, également, des mouvements discrets des commissures labiales; le rythme cardiaque s'accélère, les variations respiratoires sont très nettes, car cette malade présente, à l'état de veille, une respiration de type périodique, qui laisse place, durant la P. P., à une respiration ample et continue. Ce phénomène était si caractéristique qu'il était possible, uniquement par le bruit de la respiration à travers l'orifice de trachéotomie, de repérer à quels moments apparaissaient les phases paradoxales.
C.- Discussion
L'ensemble des faits recueillis chez le chat et chez l'homme permet d'admettre que l'on peut opposer également chez l'homme, malgré des aspects de sommeil fort différents du chat, deux stades opposés:
1 - D'une part, les stades 2, 3 et 4 (caractérisés par la présence de fuseaux et d'ondes lentes) qui sont totalement absents chez notre sujet décortiqué, doivent être rapprochés du stade de fuseaux et d'ondes lentes observé chez le chat. Les expériences pratiquées chez le chat ne laissent aucun doute quant à l'importance capItal.e du néocortex dans le déterminisme de tels éléments de synchronisation ou d'ondes lentes. Les ondes lentes nécessitent donc, de façon absolue, la présence du néocortex.
2 - La similarité, en tous points remarquable, des phénomènes périphériques, au cours de la P. P. du sommeil chez ]e chat et le stade du sommeil, accompagné de mouvements des yeux chez l'homme, permet d'admettre qu'il existe également chez celui-ci un stade dépendant de la mise en jeu du tronc cérébral inférieur.
Nous avons vu que; chez le chat, l'activité rapide corticale dépendait de mécanismes différents de ceux de l'éveil et probablement d'un circuit rhinencéphalique; il est possible que l'activité lente, en "dents de scie", recueillie chez l'homme dans les régions antérieures, puisse traduire la mise en jeu d'un tel système rhinencéphalique.
La signification psychologique de la P. P pose un problème extrêmement important, que nous nous réservons de discuter ultérieurement. Si nous n'avons pas retrouvé, avec la même fréquence que Kleitman, des souvenirs de rêves au cours de la P. P. (nous avons observé un nombre beau coup moins grand de sujets que celui-ci), les rapports de la phase paradoxale et du rêve semblent néanmoins certains. Si l'on peut transposer les résultats acquis chez les chats à l'homme, on doit alors admettre que l'activité onirique dépend d'un système rostral axé sur le tronc cérébral inférieur, probablement du au "limbic mid brain circuit". Ceci confirmerait les hypothèses qui font du rhinencéphale un circuit important dans certains phénomènes de mémorisation.
Dement et Kleitman (1957) admettent que les mouvements des yeux observés au cours de la P. P. correspondent à ´l'imagerie visuelle ª du rêve. Cependant, nous avons obtenu des mouvements des yeux au cours de la P. P. chez des animaux décortiqués chroniques ou sectionnés à la jonction méso-diencéphalique et montré qu'ils dépendaient de la partie antérieure du tronc cérébral. D'autre part, il est surprenant d'observer ces mouvements des yeux, en tous points identiques à l'individu normal, chez une femme ´ inconsciente ª depuis plus de trois ans et demi. Pour cette raison, nous pensons que des vérifications sont encore nécessaires avant d'affirmer que ces mouvements traduisent la perception onirique d'images visuelles.
Nous pensons, enfin, qu'il est illusoire de tenter de classer la P. P. par rapport aux autres stades du sommeil (1, 2, 3, 4), car il s'agit d'un phénomène qualitativement; différent, très vraisemblablement neuro-humoral qui peut apparaître périodiquement au cours de n'importe quel stade de sommeil.
Conclusions
Deux systèmes différents entrent en jeu au cours du sommeil physiologique chez le chat et chez l'homme:
1 - Le premier système se traduit par des phénomènes de synchronisation et des ondes lentes corticales et sous corticales (stade de fuseaux et d'ondes lentes du chat. Stades 2, 3 et 4 chez l'homme). Il nécessite la présence du néocortex et traduit l'action inhibitrice de celui-ci sur le système réticulé activateur ascendant. Il peut être mis en jeu par la répétition de stimuli dans le cas d'inhibition supra-liminale de type pavlovien ou par conditionnement. Il peut, également, être mis en jeu par stimulations à basse fréquence du thalamus chez l'animal; ce stade de sommeil télencéphalique représenterait un stade acquis au cours de la télencéphalisation et pourrait ainsi être qualifié de "néo sommeil".
Ce stade, au cours duquel les phénomènes d'inhibition envahissent le cortex et le sous-cortex, s'accompagne d'une absence totale d'activité ´ subjective cérébrale. Tout réveil au cours de ce stade est dénué de souvenirs oniriques.
2 - Le deuxième système est responsable de la phase paradoxale du sommeil (activité rapide corticale, comparable à celle de l'éveil avec diminution des réponses évoquées chez le chat. Activité de bas voltage sans fuseaux ni K complexes chez l'homme).
Ce stade s'accompagne constamment de phénomènes somato-végétatifs particuliers et fort significatifs (mouvements rapides des yeux, diminution totale du tonus musculaire, variations respiratoires et cardiaques). Il traduit la mise en jeu périodique d'un système axé sur la partie caudale du tronc cérébral (F. R. mésencéphalique postérieure et F. R. pontique antérieure), en relation probable rostralement avec un circuit rhinencéphalique (limbic mid brain circuit) et en relation caudale avec la F. R. inhibitrice.
Ce stade de sommeil existe seul chez l'animal mésencéphalique chronique. Il représenterait ainsi l'expression d'un archéo-sommeil. Il est plus profond que le premier stade, car le seuil d'éveil est augmenté par rapport à la phase d'ondes lentes du sommeil. Il peut être déclenché chez l'animal par stimulation du tronc cérébral inférieur. Il s'accompagne, dans un nombre très important de cas chez l'homme, d'une activité onirique. Les causes de sa mise en jeu périodique restent encore à être déterminées, mais certains résultats expérimentaux sont en faveur d'un mécanisme neuro-humoral.
Rappel méthodologique
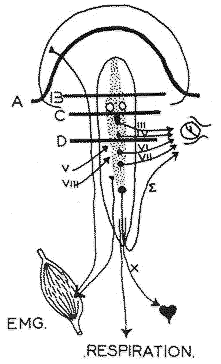
Le sommeil est une activité totale de l'organisme, jugée non seulement sur l'activité électrique corticale mais sur les corrélations de l'activité électrique sous-corticale (A: animal décortiqué; B: a. mésencéphalique; C: a. pontique; D: a. médiopontique) et les efférences somato-végétatives: pupilles, membranes nictitantes, paupières, tonus musculaire, rythme respiratoire et cardiaque.
Eveil - Sommeil lent - P.P.
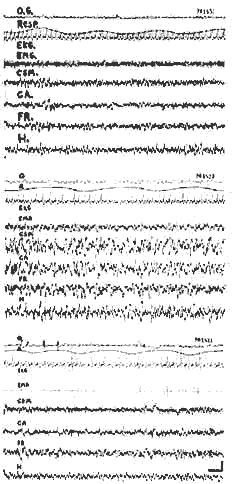
- Eveil: activité rapide corticale et sous-corticale, activité E.M.G importante au niveau des muscles de la nuque
- Sommeil lent: ondes lentes corticales et sous-corticales diminution discrète de l'activité E.M.G.; ralentissement respiratoire (Resp.).
- P.P.: activité rapide corticale et sous-corticale identique à celle de l'éveil; mais disparition totale de l'activité E.M.G., irrégularité respiratoire mouvements rapides des yeux (O.G.), ralentissement du rythme cardiaque. - C.S.M.C.A.: Cortex sensori moteur et acoustique. - F. R.: F, R. mésencéphalique. - H.: Hippocampe ventral, - Cal: 1 sec. 50 microvolts.
Eveil - Phase télencéphalique - Phase paradoxale
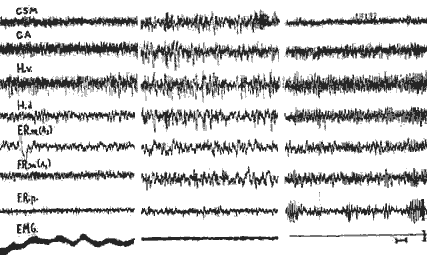
I) Eveil;
II) Phase télencéphalique du sommeil;
III) Phase paradoxale, apparition de fuseaux au niveau de la F.R. pontique F.R.P.), activité rythmique au niveau de l'hippocampe dorsale, et disparition de toute activité E.M.G, - Hv, Hd: Hippocampe ventrale, dorsale, - F. R, m.: F. R. mésencéphalique, - C.S,M.C.A.: Cortex sensori moteur et acoustique,- Noter la vitesse réduite d'enregistrement (7,5 m,/sec.), Cal: 1 sec. 50 microvolts.
Phase paradoxale
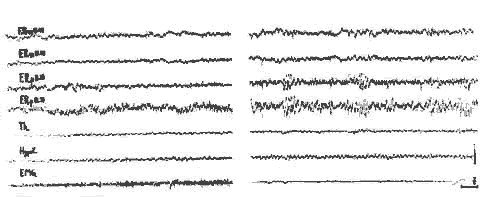
Phase paradoxale (rhombencéphalique) du sommeil chez un chat décortiqué chronique
I. - Eveil.
II. - Apparition de fuseaux au niveau du nucleus pontic caudalis (F.R.P.) et disparition du tonus musculaire (E.M.G.). Vitesse d enregistrement: 7,5 m./sec. Cal: 1 sec.
- F. R. m: F.R. mésencéphalique.
- Th.: thalamus médian
- Hypo T: Hypothalamus.
Animal mésencéphalique chronique
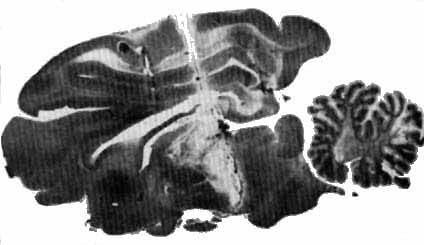
Animal mésencéphalique chronique (survie de dix jours): La section totale passe à la jonction entre le mésencéphale et le diencéphale.
Veille - P.P.
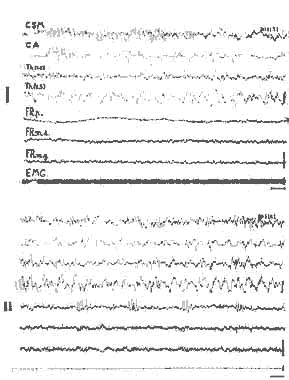
(même animal que la fig. 5). ETAT DE VEILLE: en avant de 10 section, l'activité du cortex moteur et acoustique (C.S.M.C.A.) et du thalamus (th) est lente avec des fuseaux; en arrière de la section l'activité reste rapide, car les ondes lentes (d'origine télencéphalique ne peuvent plus envahir la formation réticulée mésencéphalique (F.R.M.) ou pontique (F.R.P.). Activité musculaire importante (E.M.G.).
II. - P. P. (ARCHÉO-SOMMEIL): en avant de la section, l'activité reste identique car les voies ascendantes responsables de l'activité rapide sont sectionnées; en arrière de la section, apparaissent les signes caractéristiques de la P. P.: fuseaux au niveau de la F. R. pontique disparition totale de l'activité E.M.G. (cette P.P. a duré treize minutes). Cal: 1 seconde; 50 microvolts.
Sections du tronc cérébral
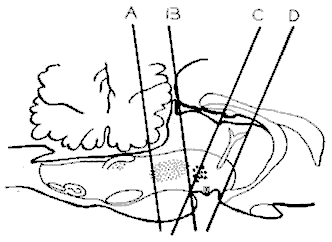
Représentation schématique des différentes sections du tronc cérébral mettant en évidence le système responsable de la P.P. :
- D) Section à la limite antérieure du mésencéphale. Une telle préparation possède tous les signes somato-végétatifs de la P. P.
- C) Section à la partie moyenne du mésencéphale (au niveau du noyau rouge). Il n'y a plus de mouvements des yeux au cours de la P.P. mais il existe toujours des variations cardio-respiratoires et une disparition du tonus musculaire.
- B) Préparation rostro-pontique: identique à la précédente.
- A) Préparation médio-pontique (Midpontine pretrigeminal preparation). Il n'existe plus de variations périodiques cardio-respiratoires ou du tonus musculaire. Le système responsable de la P.P. se situe donc entre A et B, c'est-à-dire au niveau de la F.R. pontique antérieure et mésencéphalique postérieure. Le cervelet n'est pas en cause puisque des P.P. ont été enregistrées sur des chats totalement cérébellectomisés.
Lésion tegmentum mésencéphalique
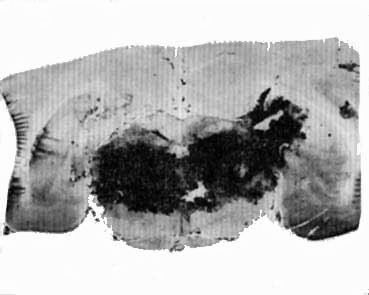
Chat W2: Lésion expérimentale intéressant totalement le tegmentum mésencéphalique, détruisant la F.R., mais respectant la partie ventrale du mésencéphale (en particulier le noyau interpédonculaire).
Eveil - P.P.
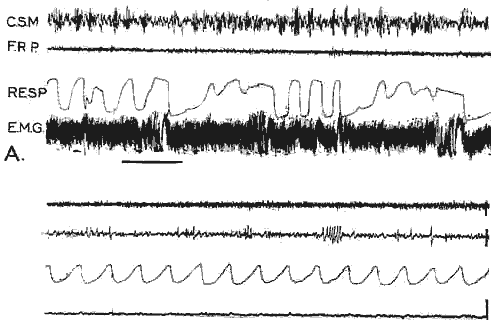
Chat W2 (voir figure précédente) :
- a) Eveil: le trait noir indique une stimulation nociceptive de la queue qui entraîne une réaction musculaire et respiratoire. Il n'y a pas d'activation corticale (C.S.M.) car le système réticulaire activateur ascendant est interrompu.
- b) P.P.: disparition totale de l'activité E.M.G., apparition de fuseaux au niveau de la F.R. pontique (F.R.P.), et activité rapide corticale.
- Cal: 2 sec. 50 microvolts
- Les structures nerveuses responsables de la P.P. sont donc différentes de celles de l'éveil
Déclenchement de la P.P. chez un chat intact
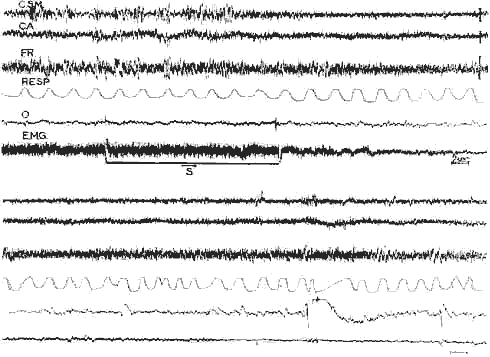
Déclenchement de la P.P. chez un chat intact par stimulation du tronc cérébral: Le, tracés I et II se font suite, I) Sommeil télencéphalique avec fuseaux corticaux et réticulaires mésencéphaliques (F.R.), Le trait horizontal signale la stimulation de la F.R, pontique antérieure (pendant 12 secondes, 0,8 Volts 300 sec., 1 millisec.)
Apparition d'une activité rapide corticale, d'une disparition progressive de l activité E.M,G. des muscles de la nuque. des mouvements rapides des yeux (O) et d'irrégularité respiratoire.
Cette P. P. a duré 10 minutes. - Cal: 2 sec. 50 microvolts
Déclenchement de la P.P. chez l'animal mésencéphalique
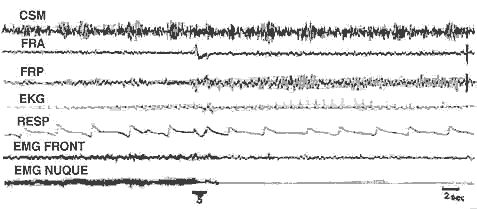
Déclenchement de la P.P. chez l'animal mésencéphalique: Le trait noir (S) indique une stimulation de la F.R pontique (F.R.P.) pendant 1 seconde (300/sec., 1 volt) Disparition totale du tonus musculaire (E.M.G.), apparition de fuseaux au niveau de la F.R.P. Il n'y a pas d'activation corticale (C.S.M.) car la section du tronc est totale. Cette P.P. provoquée a persisté 18 minutes.
Action de l'atropine et de l'éserine

Chat mésencéphalique chronigue(U2):
- A) P.P. spontanées périodiques. La flèche indique l'injection intra-péritonéale de 0,2 mg./kg. de sulfate d'atropine. Disparition presque complète des P.P. (schéma résumant un enregistrement continu d'une durée de neuf heures).
- B) Même animal, trois jours après A. P.P. spontanées périodiques. La flèche repère l'injection intra-péritonéale de 0,02 mg./kg. de sulfate de physostigmine (esérine) La durée des P P. est augmentée.
Structures mises en jeu au cours du sommeil
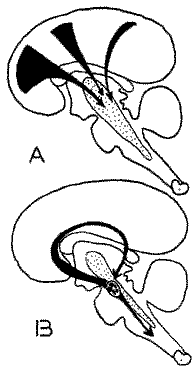
Représentation très schématique des structures mises en jeu au cours du sommeil:
- a) inhibition corticifuge envahissant la formation réticulée mésencéphalique (sommeil avec fuseaux et ondes lentes);
- b) phase paradoxale du sommeil. Mise en jeu d'un mécanisme situé au niveau du tronc cérébral inférieur. en relation avec le limbic midbrain circuit en avant, et avec la F.R. inhibitrice en arrière.
Aspects E.E.G. du sommeil chez l'homme
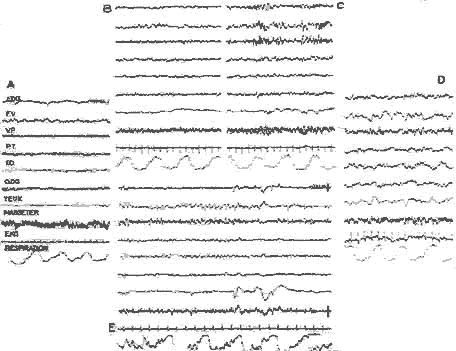
- a) Eveil, avec rythme alpha;
- b) Stade 1: activité rapide de bas voltage, à l'endormissement;
- c) Stade 2: fuseaux;
- d) Stade 4: ondes lentes;
- e) P.P.: activité de bas voltage, ondes lentes rythmiques antérieures, "en dents de scie", mouvements des yeux, irrégularité respiratoire. - Ralentissement du pouls.
Détail de la P.P.
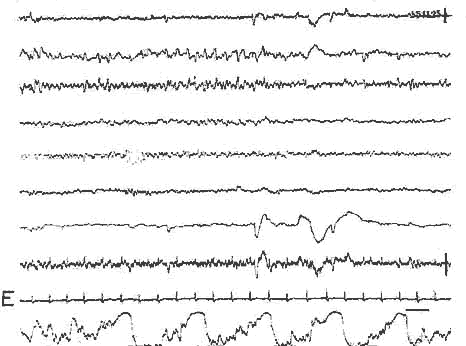
Périodicité de la P.P.
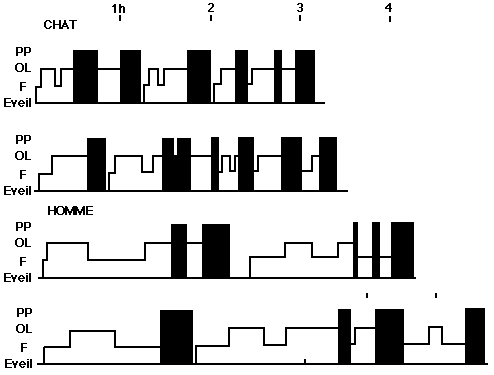
Périodicité de la P.P. chez le chat et chez l'homme: Représentation schématique des niveaux de sommeil au cours de deux périodes de sommeil chez le chat et de deux nuits chez l'homme (de minuit à 4 et 5 heures du matin). Les périodes d'éveil, de fuseaux (F.) et d'ondes lentes (O. L.) sont représentées en blanc, les P. P. en noir. Echelle de temps en heure.
Variations cardio-respiratoires
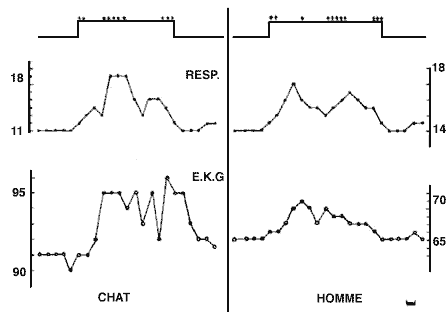
Variations cardio-respiratoires au cours de la phase paradoxale, chez un chat intact et un sujet adulte. Les croix indiquent les mouvements des yeux. Echelle de temps: 1 minute. Durée de la P.P.: chat: 10 minutes; homme: 15 minutes.
Ventriculographie
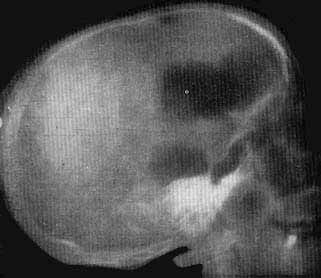
Dilatation importante des ventricules.
Périodicité de la P.P.
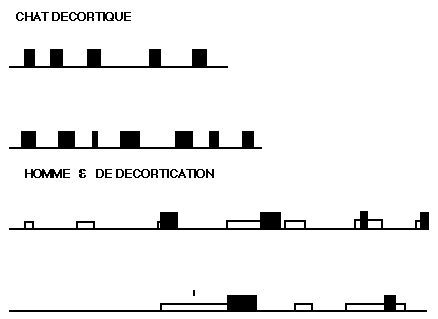
Périodicité de la P.P. chez un chat décortiqué, et chez une malade présentant un syndrome de décortication: Les P.P. sont représentées en noir. Chez l'homme, les pointillés marquent des périodes de sommeil comportemental avec disparition de l'hypertonie musculaire sans mouvements des yeux (voir fig. 19).
Veille - Endormissement - Activité rapide
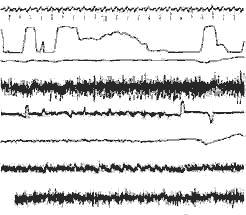
I) Veille:
Activité rapide au niveau du scalp, masquée par des artéfacts
musculaires. Intense activité E. M. G. au niveau des fléchisseurs
(hypertonie).
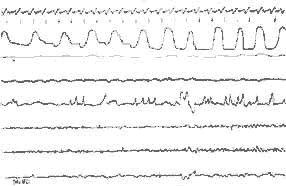
II) Endormissement:
Le tracé E.E.G. ne varie pas, disparition de l'activité
musculaire, absence de mouvements des yeux.
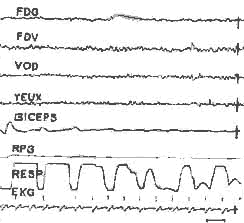
III) P. P.: Activité rapide avec quelques ondes lentes
rythmiques au niveau du vertex. Mouvement des yeux, accélération
respiratoire. Ralentissement important du rythme cardiaque, et disparition
totale de l'activité E.M.G. des fléchisseurs.