| Eveil, sommeil, rêve |
| Michel Jouvet Le courrier du CNRS |
Notre cerveau subit l'alternance de trois états de vigilance principaux: l'éveil, le sommeil et le rêve. Si les fonctions du sommeil et de l'éveil sont connues, en revanche la fonction de ce troisième état du cerveau qu'est le rêve demeure l'une des énigmes les plus irritantes de la biologie.
L'alternance activité-repos appartient au monde vivant végétal ou animal et nous semble ainsi naturelle. L'éveil est nécessaire à notre survie et s'accompagne des dépenses d'énergie demandées par la satisfaction des besoins immédiats de l'individu ou de l'espèce. Deux notions intuitives expliquent le sommeil: la première est écologique et rend compte d'un héritage génétique que nous tenons de nos ancêtres, les premiers vertébrés à sang chaud (Homéothermes). Comme ils étaient placés dans des conditions où la nourriture leur était insuffisante, l'évolution a d'abord "inventé" l'hibernation, puis la torpeur. Ainsi, la diminution de température centrale permettrait une économie d'énergie et une survie prolongée en l'absence de nourriture. Le sommeil serait l'héritier de ces mécanismes. La seconde notion est fondée sur le concept subjectif de fatigue: l'activité cérébrale ou musculaire entraîne la fatigue et nécessite donc une période de "repos" cérébral pendant lequel les cellules nerveuses pourraient devenir inactives. Activité-repos, éveil-sommeil apparaissent ainsi rentrer dans un cadre naturel logique. En fait, ce schéma est trop simple et ne tient pas compte de la réalité. Il faut admettre, d'une part, que notre cerveau (comme celui de tous les Homéothermes: des oiseaux à l'homme) subit l'alternance non de deux mais de trois états de vigilance principaux: l'éveil, le sommeil et le rêve. D'autre part, nous savons que le sommeil n'est pas un repos du cerveau, stricto sensu, mais un phénomène actif puisque celui-ci consomme au moins autant de glucose et d'oxygène que pendant l'éveil.
L'éveil et ses degrés
L'aventure de l'homme pourrait être celle de la conquête de l'éveil. La tendance naturelle de l'homme est, en effet, de s'endormir spontanément si aucune tâche physique ou intellectuelle ne lui est proposée. C'est pour conquérir un éveil plus durable et plus efficient que l'homme a découvert le thé, le café qui augmentent l'efficacité du système d'éveil. Ce système d'éveil est situé au niveau du tronc cérébral (dans la formation réticulaire du mésencéphale): les cellules réticulaires reçoivent, par l'intermédiaire de collatérales des voies sensorielles, des informations du milieu extérieur. Ces informations peuvent venir converger sur un seul neurone. Elles perdent alors leur signe spécifique d'origine pour acquérir la propriété non spécifique de mettre en jeu le réseau de neurones qui va venir exciter le cortex cérébral et déclencher la réaction d'éveil qui se caractérise par une accélération du rythme de l'activité électrique du cortex cérébral. D'autres cellules réticulaires augmentent le tonus musculaire. Ainsi, les mécanismes de base de l'éveil sont effectués par ce système nécessaire, mais non suffisant, pour l'attention, l'apprentissage ou l'exécution d'un geste. En effet, si on enlève le cortex cérébral d'un animal, il conserve la possibilité d'être éveillé, mais ne peut apprendre. On admet actuellement que la formation réticulaire mésencéphalique accélère l'activité corticale par l'intermédiaire d'un neuro transmetteur, l'acétylcholine. Celle ci est contrôlée au niveau réticulaire et cortical par un autre médiateur, la noradrénaline. La dopamine joue également un rôle dans la facilitation de l'initiation des mouvements au cours de l'éveil. L'expérience suivante illustre le rôle éveillant de ces deux neurotransmetteurs: on sait que les amphétamines provoquent un éveil intense et durable en augmentant la libération de la noradrénaline et de la dopamine. Si l'on empêche la synthèse de ces deux neurotransmetteurs (en inhibant une enzyme de leur chaîne de synthèse), l'injection secondaire d'amphétamine n'a alors plus aucune action éveillante. La caféine possède une action éveillante différente des amphétamines car elle peut agir directement sur le cortex cérébral. Aussi, son action éveillante se double-t-elle d'une action psychoto nique bien connue.
Récemment, un système de neurones histaminergiques a été découvert dans l'hypothalamus ventrolatéral. Il semble également jouer un rôle important dans l'éveil (ce qui explique que des drogues anti-antihistamines aient un pouvoir "anti-éveil" bien connu).
Acétylcholine, catécholamines, sérotonines (voir plus loin), histamine ne sont pas les seuls "médiateurs" de l'éveil. Il faut leur ajouter certains peptides comme le TRH ou le MIF dont les mécanismes intimes d'action sont encore inconnus. En fait, l'éveil est tellement important pour la survie de l'espèce qu'il est fort probable que de nombreux systèmes, plus ou moins redondants, ont été acquis successivement. Ils entrent en jeu pour assurer les mécanismes complémentaires survenant au cours de l'éveil: faim, soif, activité sexuelle, apprentissage, mémoire, etc.
S'il est possible d'augmenter la qualité de l'éveil par des drogues, il est plus difficile d'en augmenter la durée sans se heurter à des phénomènes d'accoutumance (nécessité de doses plus grandes), d'assuétude (dépendance du sujet vis-à-vis de la drogue) ou de toxicité (de fortes doses d'amphétamines provoquent un tableau de psychose aiguë). Enfin, tout éveil prolongé est suivi d'une augmentation secondaire du sommeil, car c'est pendant l'éveil que nous produisons les facteurs responsables du sommeil.
Le sommeil
Diminution réversible naturelle et périodique de la perceptivité du milieu extérieur avec conservation d'une réactivité et conservation des fonctions végétatives. Cette vieille définition reste valable car elle permet de distinguer le sommeil du coma: un bruit violent réveille un dormeur mais pas un comateux. Depuis vingt ans, grâce à la polygraphie, la neurochimie et la polarographie, des progrès considérables ont été accomplis dans la connaissance du sommeil. Nous savons ainsi que le sommeil n'est pas un processus homogène, mais qu'il s'agit de la juxtaposition périodique de deux états: le sommeil proprement dit et le sommeil paradoxal (qui correspond au rêve et que nous étudierons plus loin).
Chez l'homme, les différents stades du sommeil, selon la "profondeur", c'est-à-dire l'intensité d'un stimulus capable de réveiller un dormeur, s'enchaînent dans le temps, depuis le stade I ou sommeil léger de l'endormissement, jusqu'aux stades II, III et IV caractérisés par le ralentissement de la fréquence de l'activité électrique cérébrale d'où le terme de "sommeil lent". Le sommeil s'accompagne de concommitants végétatifs; par exemple, la tension artérielle et le rythme cardiaque diminuent ainsi que la température centrale; la respiration également qui peut s'accompagner de ronflements. Les modifications respiratoires du sommeil sont parfois dramatiques chez le nourrisson puisque les commandes végétatives des centres respiratoires peuvent alors se dérégler (mort subite du nourrisson). On sait également que les stades III et IV du sommeil commandent la sécrétion d'hormones de croissance. Ce phénomène est important chez l'enfant puisqu'on a pu mettre sur le compte d'un mauvais sommeil dû à des conditions socio-économiques mauvaises (bruit, promiscuité) certains retards de croissance qui cèdent si l'enfant peut dormir correctement.
Le sommeil n'est pas la conséquence d'un repos passif du système d'éveil mais il est provoqué par l'intervention d'un mécanisme actif. L'activité des neurones corticaux ne s'arrête pas au cours du sommeil mais subit des modifications particulières (synchronisation) que l'on a pu interpréter comme la mise en marche de circuits empêchant l'intégration des signaux du monde extérieur. Les mécanismes de l'endormissement et du sommeil ne sont sans doute pas univoques. On s'endort au cours d'une conférence ennuyeuse, après un bon repas ou si l'on reste trop longtemps éveillé. Deux théories complémentaires expliquent actuellement de nombreux faits, mais pas tous. Selon la première, qui remonte à Piéron, il y aurait accumulation dans le liquide céphalo-rachidien, au cours de l'éveil, de substances hypnogènes (appelées au début hypnotoxines). On pense actuellement qu'il pourrait s'agir de peptides d'origine hypothalamique qui pourraient inhiber les neurones du système d'éveil (l'injection du liquide céphalo-rachidien d'un animal donneur, privé de sommeil, dans le système ventriculaire d'un animal receveur peut augmenter la profondeur et la durée de son sommeil). Un neuromédiateur libéré au cours de l'éveil, la sérotonine, synthétisée par des cellules situées sur la ligne médiane du tronc cérébral (le système du Raphé) serait responsable du sommeil. La destruction des cellules qui synthétisent la sérotonine au niveau du système du Raphé entraîne, en effet, une insomnie de très longue durée (plusieurs jours). D'autre part, le blocage de la synthèse de la sérotonine entraîne aussi une insomnie - qui peut être réversible en sommeil physiologique si l'on injecte le précurseur immédiat de la sérotonine. Il est possible que la sérotonine agisse comme une véritable hormone cérébrale. En pénétrant dans certains neurones, elle déclencherait la synthèse de peptide hypnogène. L'action endormante du précurseur de la sérotonine peut, en effet, être supprimée par blocage de la synthèse des protéines. L'incertitude concernant les mécanismes du sommeil empêche la réalisation de véritables drogues hypnogènes physiologiques. Alors que l'homme a su trouver dans la nature des produits augmentant l'éveil, il n'a pas encore trouvé de molécules hypnogènes naturelles. Les opiacés et leurs dérivés ne sont pas de véritables hypnotiques. Les barbituriques entraînent le sommeil en déprimant l'activité du système d'éveil, mais leur excès entraîne une narcose ou un coma. Les benzodiazépines sont moins dangereuses, mais ont l'inconvénient de diminuer le rêve.
Le sommeil paradoxal
La dualité des états du sommeil avait été déjà devinée par les mystiques Hindous qui opposaient au sommeil sans rêve (Prajna), l'éveil intérieur du rêve (Taijasa). Cependant, l'activité onirique ne fait son entrée en physiologie comme troisième état du cerveau il y a une vingtaine d'années seulement. Comme le montre l'enregistrement polygraphique du sommeil (hypnogramme), 4 à 5 périodes de sommeil paradoxal, d'une durée de 15 à 20 minutes (soit environ 100 minutes, soit 20 % de la durée totale du sommeil), surviennent chaque nuit. Elles sont caractérisées par une accélération du tracé cortical (qui redevient similaire au stade I de l'endormissement). En fait, il ne s'agit pas du retour du stade I de sommeil léger, car le sommeil paradoxal est aussi "profond" que les stades III et IV. D'autre part, une constellation de signes périphériques vient affirmer qu'il s'agit d'un autre état: apparition de mouvements oculaires rapides, qui contrastent avec l'atonie musculaire totale (ce qui explique l'absence de réflexes tendineux à ce moment, ou la chute brusque de la tête d'un individu qui dort assis). La respiration devient irrégulière, la tension artérielle subit des variations brusques (qui peuvent expliquer les accidents vasculaires, ramolissements. ou infarctus survenant au cours du sommeil). Il existe enfin une érection, manifestation végétative dont le contenu sexuel est peu probable puisqu'on l'observe aussi bien chez le nouveau-né que chez le vieillard. Réveillé au cours d'une période de mouvements oculaires, le dormeur racontera en détail des souvenirs de rêve, alors que réveillé à d'autres moments du sommeil, ces souvenirs sont soit absents,soit plus estompés et surtout ont perdu le caractère fantastique de l'imagerie onirique.
La découverte de ce troisième état du cerveau qu'est le rêve ou sommeil paradoxal chez l'homme et les mammifères a plongé les neurophysiologistes dans un grand étonnement. L'alternance veille-sommeil peut s'expliquer naturellement, mais pourquoi un troisième état qui semble traduire un besoin ? En effet, si l'on supprime le rêve (en réveillant un dormeur au début du sommeil paradoxal), celui-ci tend à revenir de plus en plus souvent. Il existe aussi, après cessation des privations pharmacologiques de rêve (par exemple avec les hypnotiques ou les inhibiteurs des monoamines-oxydases) une augmentation considérable du sommeil paradoxal qui peut alors survenir directement au cours de l'éveil. Ce qui se traduit par la narcolepsie (ou maladie de Gélineau) avec effondrement cataplectique (perte subite du tonus musculaire et brève période d'activité onirique).
L'exploration du sommeil paradoxal (ce nouveau continent du cerveau) est loin d'être terminée, mais la moisson de résultats obtenus depuis vingt ans peut être résumée ainsi: il semble que le sommeil paradoxal apparaisse au cours de l'évolution avec l'acquisition de l'homéothermie (c'est-à-dire avec les oiseaux et les mammifères) Il n'a pas été possible, en effet, de déceler le sommeil paradoxal chez les poissons, les amphibiens ou les reptiles, qui sont des Poïkilothermes. Le sommeil paradoxal est donc une "acquisition phylogénétique" relativement récente. En revanche, c'est un phénomène ontogénétiquement précoce. L'étude des embryons d'oiseaux in ovo, des foetus in utero semble bien montrer que le sommeil paradoxal constitue la majeure partie, sinon la totalité du sommeil au cours de la maturation du système nerveux. Il existe ensuite, après le sevrage, des différences considérables entre les différentes espèces qui peuvent être expliquées en partie par des considérations éco-éthologiques. Le sommeil paradoxal dépend étroitement du sommeil profond qui le précède.
Le facteur "sécurité" joue alors un grand rôle (l'insécurité entretenant les mécanismes de l'éveil et empêchant le sommeil) ainsi que les conditions de vie. L'herbivore doit passer beaucoup de temps éveillé pour absorber une nourriture peu énergétique, ce qui n'est pas le cas des carnivores. Ainsi, la durée quotidienne du sommeil paradoxal, qui n'est que de 10 à 15 minutes chez la vache, atteint 180 à 200 minutes chez le chat domestique.
Mécanismes du sommeil paradoxal
Bien que lié étroitement au sommeil lent, le sommeil paradoxal dépend de structures nerveuses différentes. Trës schématiquement, on peut admettre que le sommeil paradoxal met en jeu deux systèmes différents dont le point de départ se situe dans la formation réticulée du pont (au-dessous du mésencéphale). Le premier système peut être comparé à un "générateur" endogène qui met en jeu les mouvements oculaires et vient exciter la grande majorité des neurones cérébraux, que ce soit des aires sensorielles ou motrices. Cependant, mis à part les mouvements oculaires, la mise en jeu des systèmes moteurs cérébraux n'entraîne pas de mouvements globaux des membres ou du corps, car il existe un deuxième système responsable de l'atonie qui vient bloquer l'excitation de la voie finale motrice commune au niveau de la moelle; c'est pourquoi le somnambulisme ne correspond pas à l'activité onirique mais traduit un éveil incomplet au cours du sommeil lent. La destruction chez le chat du système responsable de l'atonie a permis de dévoiler l'apparition de "comportements oniriques" stéréotypés de jeu, de chasse, d'attaque et de rage contre d'imaginaires partenaires au cours du sommeil paradoxal.
Les mécanismes biochimiques du rêve sont complexes, car l'apparition du rêve est inhibée à la fois par le système d'éveil et certains systèmes mis en jeu au cours du sommeil. On pense actuellement que des facteurs situés dans les systèmes hypothalamo-hypophysaires joueraient un rôle décisif dans le déclenchement du sommeil paradoxal. L'action de ce (ou ces) facteur(s) et la levée de l'inhibition due aux neurones à sérotonine viennent libérer les "mécanismes exécutifs" du rêve au sein desquels l'acétylcholine joue un rôle primordial. Toutes ces étapes complexes rendent le rêve très susceptible à de nombreuses drogues qui sont capables de le supprimer, alors qu'on ne connaît pas (chez l'homme) de drogues capables de le provoquer bien qu'on puisse le faire chez le chat par injection intracérébrale de drogues agissant sur l'acétylcholine, ou par injection intraventriculaire des facteurs hypothalamiques.
Horloge biologique et cycle éveil-sommeil-rêve
Le cycle éveil-sommeil, chez la plupart des espèces, est sous la dépendance du rythme nycthéméral (le jour et la nuit). Ce rythme agit comme synchroniseur ou donneur de temps (Zeitgeber) sur une horloge biologique interne située dans l'hypothalamus, au niveau des noyaux suprachiasmatiques qui reçoivent l'information lumineuse par des voies venant de la rétine. Cette horloge biologique interne, en l'absence de synchroniseur externe (par exemple un séjour prolongé dans un abri ou une grotte) fonctionne "en libre cours" avec un petit retard ou une avance sur le rythme de 24 h, donc à peu près 24 h (circadies), d'où le terme de rythme circadien. Or, cette horloge interne tient sous sa dépendance de nombreuses autres horloges qui contrôlent la synthèse d'enzymes et d'hormones, la température centrale et indirectement le rythme d'éveil et de sommeil. Cela n'a pas eu d'importance autrefois, mais la civilisation industrielle avec le travail posté (rythme des 3 x 8), les horaires irré guliers (les routiers), et surtout l'apparition des Jets et des voyages intercontinentaux, ont placé le cerveau humain dans des conditions que l'évolution n'avait pas prévues. Ainsi, un voyageur quittant Lyon pour Los Angeles conserve le rythme circadien endogène de son horloge interne, alors qu'il reçoit les informations jour-nuit décalées de 8 ou 9 heures. Il en résulte une inadéquation (désynchronisation) entre son besoin de sommeil ou d'éveil et le temps légal (d'où réveil au milieu de la nuit et endormissement le jour). Cette désynchronisation entre l'horloge circadienne endogène et le temps réel peut entraîner, surtout si elle est répétée souvent, des dérèglements du cycle éveil-sommeil (endormissement invincible) et parfois des troubles psychiatriques (dépression). Si le rythme circadien commande l'alternance veille-sommeil, c'est un rythme ultradien de l'ordre de 90 minutes qui règle la périodicité du sommeil paradoxal. Chez l'homme, il est possible que ce rythme ultradien soit l'expression d'un rythme fondamental (Basic Rest Activity "BRAC"). Ce rythme serait alors prédominant chez le nouveau-né, mais serait ensuite masqué au cours de l'éveil chez l'adulte. Il réapparaîtrait alors au cours de la narcolepsie dont les accès diurnes ont souvent une périodicité de 90 minutes. On conçoit ainsi combien apparaît important, en ergonomie et en clinique, de surveiller les possibles asynchronies entraînées par les horaires alternants ou les répétitions de voyages intercontinentaux.
Les fonctions des états de vigilance
Si les fonctions de l'éveil sont la condition nécessaire de la survie de l'individu et de l'espèce, les fonctions du sommeil et surtout du rêve demeurent l'une des énigmes les plus irritantes de la neurobiologie. Il est possible que le sommeil soit l'aboutissement actuel des mécanismes de conservation de l'énergie, mais les retentissements hormonaux du sommeil lent font supposer que des processus de synthèse protéique peuvent également survenir et jouer sans doute un rôle à la fois au niveau du système nerveux central (phénomènes de mémoire à long terme) et de l'organisation (nanisme par déficit de sommeil). Cependant, l'existence de sujets en très bonne santé n'ayant dormi que 3 à 4 h par nuit pendant des décennies permet de supposer que deux cycles de sommeil (180 minutes) pourraient être suffisants et qu'il existe un quota nécessaire et un quota de luxe dans notre sommeil.
L'autre fonction du sommeil est de préparer et d'ouvrir les portes du rêve: mais les fonctions supposées de l'activité onirique sont aussi nombreuses que les chercheurs qui l'étudient. Il est difficile de supprimer sélectivement le rêve sans provoquer des effets non spécifiques dus au "stress" de la méthode instrumentale ou dus à la multiplicité d'action des diverses drogues qui suppriment le sommeil paradoxal. D'autre part, on sait maintenant que la plupart des méthodes instrumentales ou pharmacologiques supprimant le sommeil paradoxal provoquent une augmentation du ou des "facteurs onirogènes". Les troubles observés ne sont donc pas dus à la suppression du rêve mais au contraire à l'augmentation de ces facteurs. Certains résultats sont en faveur du rôle du sommeil paradoxal sur la mémoire chez le rat, mais cela n'a pas été vérifié chez l'homme. Selon une autre hypothèse, le rêve correspondrait à une programmation périodique génétique du cerveau qui serait responsable de l'entretien de l'hérédité psychologique qui constitue la partie héritable de la typologie d'un individu. Au cours de l'éveil, une grande partie des circuits nerveux de notre cerveau est influencée par l'environnement et l'apprentissage du fait de la plasticité cérébrale. Afin de conserver le programme établi au cours de la maturation cérébrale, et qui rend chaque individu différent des autres, malgré un environnement identique, on a supposé que le générateur qui entre en jeu au cours du rêve viendrait modifier périodique ment certains circuits selon une programmation où les facteurs génétiques joueraient un rôle important. En ce sens, le rêve serait le retour à la nature (un éveil génétique) qui s'opposerait, chez l'homme, à l'éveil lié à l'environnement épigénétique culturel.
Ainsi, le cycle éveil-sommeil-rêve nous fait peut-être, chaque jour, reparcourir sans que nous le devinions l'aventure de l'évolution de notre cerveau.
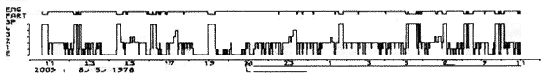
Les hypnogrammes se pratiquent habituellement pendant une durée de 24 h. Ils permettent de repérer les troubles majeurs du cycle éveil-sommeil-rêve chez certains sujets.
Ordonnées: en haut: état de l'électromyogramme (EMG) et stades du sommeil (1-2-3-4 et SP) E = éveil.
Abscisses: temps réel en heures. L = arrêt de la lumière.
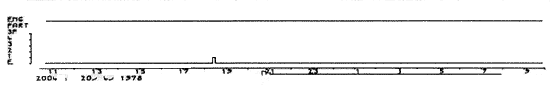
Insomnie totale due à une maladie de Morvan. Cette insomnie (appelée égulement agrypnie) a persisté pendant 2 mois.

Insomnie grave. Un seul cycle incomplet de sommeil entre minuit et 2 h et quelques minutes de sommeil léger vers la fin de la nuit. Il n'y a pas de sommeil paradoxal (Maladie dégénérative du tronc cérébral).
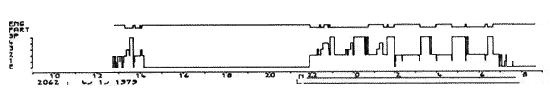
Sommeil normal. Le sommeil à l'heure de la sieste (13 h-14 h) n'est pas pathologique dans les conditions d'enregistrement.
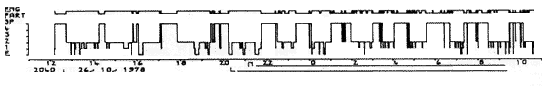
Hypersomnie considérable en SP. Le sommeil occupe la durée entière de l'enregistrement de 12 h à 10 h. Remarquer l'apparition périodique du sommeil paradoxal dont la quantité totale dépasse 300 mn (au lieu de 100 mn). Il s'agit d'un "rebond" de SP après arrêt d'un traitement de longue durée par les inhibiteurs des monoamines oxydases (qui suppriment le SP).
Narcolepsie (Maladie de Gélineau). Les épisodes de SP surviennent directement au cours de l'éveil. Ils sont invincibles et peuvent être responsables d'accidents du travail ou de la circulation. La nuit de sommeil n'est pas normale (absence de stades III et IV). Des recherches récentes chez l'animal ont permis de mettre au point des drogues de plus en plus efficaces permettant de contrôler les accès cataplectiques de la maladie de Gélineau.

L'iguane (Iguana Iguana) est bien sûr éveillé, peut dormir, mais il n'a pas encore été possible de déceler chez ce reptile des périodes de sommeil paradoxal, malgré des enregistrements prolongés pendant plusieurs mois.
Figure 1 : Insomnie totale due à une maladie de Morvan
Ordonnées: en haut: état de l'électromyogramme
(EMG) et stades du sommeil (1-2-3-4 et SP) E = éveil.
Abscisses: temps réel en heures.
L = arrêt de la lumière.
Cette insomnie (appelée égulement agrypnie) a persisté pendant 2 mois.
Figure 2 : Insomnie grave
Ordonnées: en haut: état de l'électromyogramme
(EMG) et stades du sommeil (1-2-3-4 et SP) E = éveil.
Abscisses: temps réel en heures.
L = arrêt de la lumière.
Un seul cycle incomplet de sommeil entre minuit et 2 h et quelques minutes de sommeil léger vers la fin de la nuit. Il n'y a pas de sommeil paradoxal (Maladie dégénérative du tronc cérébral).
Figure 3 : Sommeil normal
Ordonnées: en haut: état de l'électromyogramme
(EMG) et stades du sommeil (1-2-3-4 et SP) E = éveil.
Abscisses: temps réel en heures.
L = arrêt de la lumière.
Le sommeil à l'heure de la sieste (13 h-14 h) n'est pas pathologique dans les conditions d'enregistrement.
Figure 4 : Hypersomnie considérable en SP
Ordonnées: en haut: état de l'électromyogramme
(EMG) et stades du sommeil (1-2-3-4 et SP) E = éveil.
Abscisses: temps réel en heures.
L = arrêt de la lumière.
Le sommeil occupe la durée entière de l'enregistrement de 12 h à 10 h. Remarquer l'apparition périodique du sommeil paradoxal dont la quantité totale dépasse 300 mn (au lieu de 100 mn). Il s'agit d'un "rebond" de SP après arrêt d'un traitement de longue durée par les inhibiteurs des monoamines oxydases (qui suppriment le SP).
Figure 5 : Maladie de Gélineau
Ordonnées: en haut: état de l'électromyogramme
(EMG) et stades du sommeil (1-2-3-4 et SP) E = éveil.
Abscisses: temps réel en heures.
L = arrêt de la lumière.
Les épisodes de SP surviennent directement au cours de l'éveil. Ils sont invincibles et peuvent être responsables d'accidents du travail ou de la circulation. La nuit de sommeil n'est pas normale (absence de stades III et IV). Des recherches récentes chez l'animal ont permis de mettre au point des drogues de plus en plus efficaces permettant de contrôler les accès cataplectiques de la maladie de Gélineau.
Figure 6 : L'iguane
L'iguane (Iguana Iguana) est bien sûr éveillé, peut dormir, mais il n'a pas encore été possible de déceler chez ce reptile des périodes de sommeil paradoxal, malgré des enregistrements prolongés pendant plusieurs mois.