| L'histoire naturelle du rêve |
| Conférence de Michel Jouvet |
| IMPRESSION |
| Version imprimable (Tout l'article dans une seule page) |
Mécanismes du Rêve
Dans quel but l'Evolution a donc inventé le rêve chez les homéothermes ?
Un début possible d'explication est apporté par la voie réductionniste de la neurophysiologie expérimentale qui, depuis une vingtaine d'années, a réussi à démonter la machinerie intrinsèque du rêve et à répondre aux questions suivantes (fig. 22).
Quelles sont les structures cérébrales nécessaires et suffisantes au déclenchement périodique des phénomènes exécutifs du rêve ? Comment interagissent-elles ? Enfin, où se trouvent les systèmes dits permissifs qui empêchent le rêve d'apparaître au cours de l'éveil, et seulement (en général) après une phase assez longue de sommeil ?
1) Quelles sont donc les parties du cerveau suffisantes au déclenchement du rêve ?
Ni l'ablation du cortex cérébral, ni celle du cervelet, n'entraîne d'altérations significatives dans l'apparition du sommeil paradoxal. Bien plus, malgré l'ablation entière des structures situées en avant du pont (incluant l'hypothalamus et l'hypophyse), le sommeil paradoxal continue à apparaître périodiquement. Il se reconnaît par l'atonie musculaire totale, d'autant plus remarquable que l'animal dit pontique est en état de rigidité lorsqu'il est éveillé. D'autre part, les signes électriques caractéristiques du sommeil paradoxal apparaissent au niveau du pont accompagnant les mouvements latéraux des yeux et les variations cardio-respiratoires. Etant donné que le pont et le bulbe sont suffisants à l'apparition périodique du sommeil paradoxal, il doit donc se trouver au niveau de ces structures des systèmes dits exécutifs responsables à la fois de l'apparition périodique et du déroulement du sommeil paradoxal.
2) Les structures exécutives ponto-bulbaires responsables du sommeil paradoxal ou la machinerie intrinsèque primitive du rêve (fig. 23).
Les mécanismes de base du sommeil paradoxal sont responsables de deux fonctions qui sont complémentaires l'une et l'autre. D'une part, elles mettent en jeu un système endogène d'excitations du cerveau au moyen de l'activité PGO (ponto-géniculo-occipItal.e). Cette stimulation entraîne l'excitation des systèmes sensoriels (surtout visuels) et des systèmes moteurs (neurones pyramidaux de l'aire motrice). Ainsi, des influx descendants moteurs vont répondre à ces stimulations et gagner la moelle épinière pour déclencher des gestes et des comportements. C'est pour empêcher cette activité motrice qu'un deuxième mécanisme doit entrer en jeu. Il vient bloquer, par une inhibition descendante puissante, les neurones moteurs de la moelle. Ainsi, le rêveur se trouve paralysé et ne peut bouger.
a) L'activité ponto-géniculo-occipItal.e (PGO) : la topographie des neurones (fort probablement cholinergiques) qui constituent le générateur endogène de l'activité PGO du rêve a été délimitée avec précision. Elle est située dans la formation réticulée pontique. Nous connaissons également les voies qui conduisent l'activité PGO au niveau des noyaux moteurs oculaires (où elle déclenche les mouvements rapides des yeux). Les voies ascendantes menant au cortex cérébral, soit directement, soit par l'intermédiaire de relais thalamiques, ont également été délimitées avec précision. L'activité du générateur PGO semble donc intéresser (programmer ?) tout l'encéphale. C'est-à-dire que si l'on introduit une microélectrode dans n'importe quel endroit du cerveau, on a environ 60 % de chance d'enregistrer l'activité unitaire d'un neurone qui est asservie (soit augmentée, soit diminuée) par l'activité PGO du générateur. 0n ne connaît cependant pas encore sur quels types (immunohistochimiques) de cellules corticales se projette l'activité PGO, bien que l'on devine que les récepteurs mis en jeu soient nicotiniques au niveau de certains relais stratégiques.
b) Le frein moteur dépend de 3 étages. Le premier est un étage de commande qui est le seul à être soumis à une régulation par les systèmes permissifs (voir plus loin). Les autres sont des étages d'exécution. L'étage de commande bilatérale est située à côté du locus coeruleus, au niveau d'un petit groupe de cellules appelé locus coeruleus alpha. Ce groupe de cellules, dont le transmetteur est encore inconnu, est normalement freiné pendant l'éveil et le sommeil par un système permissif puissant, celui du locus coeruleus dont les terminales libèrent de la noradrénaline. Ainsi, pendant l'éveil ou le sommeil, l'activité électrique du locus coeruleus est importante, alors que celle du locus coeruleus alpha est nulle. L'activité du frein du frein (c'est-à-dire du locus coeruleus) diminue au cours du sommeil puis cesse complètement au début du sommeil paradoxal (neurones PS off). Le locus coeruleus alpha entre alors en jeu (neurones PS on) (fig. 24). L'activité électrique de ses neurones qui était silencieuse croît brusquement. Des influx excitateurs sont alors envoyés au deuxième étage, bulbaire, par l'intermédiaire d'un faisceau descendant. L'arrivée des signaux excitateurs au niveau du noyau magno-cellulaire bulbaire (appelé ainsi parce qu'il contient des cellules de grande taille) entraîne à son tour l'excitation de ce noyau qui envoie des influx descendants inhibiteurs gagnant la moelle épinière. Ils viennent bloquer, au niveau des neurones moteurs alpha (ceux qui innervent directement les muscles), l'excitation qui arrive d'autre part par le faisceau pyramidal (mis en jeu par les cellules corticales sous l'influence de l'activité PGO). Quelquefois, certains influx moteurs particulièrement puissants peuvent franchir cette barrière inhibitrice provoquant de petits mouvements des doigts, des oreilles (chez le chat) ou des vibrisses (les moustaches du chat). Seuls les neurones moteurs oculaires (et les mécanismes de la respiration) échappent à cette intense activité inhibitrice.
Représentation extrêmement schématique des processus initiant les 3 états de fonctionnement du cerveau
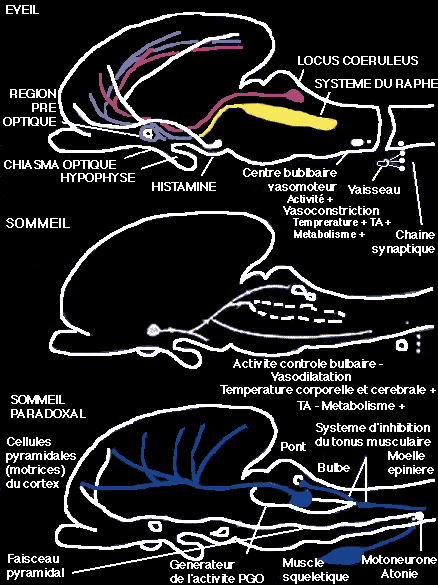
Au cours de l'éveil, il y a activation des systèmes aminergiques (sérotoninergique du raphé, noradrénergique du locus coeruleus) et histaminergique au niveau de l'hypothalamus postérieur. Un relais bulbaire adrénergique commande l'excitation du système sympathique. Il est responsable de la vasoconstriction, de l'augmentation de la température centrale et du métabolisme.
Le sommeil apparaît lorsque, sous l'influence probable de la sérotonine libérée au cours de l'éveil, la région préoptique est activée. Son activation, par l'intermédiaire d'un facteur encore inconnu, va mettre en jeu une cascade d'inhibitions intéressant les systèmes aminergiques. Il y a également diminution de l'activité du système bulbaire, vasodilatation, donc perte de chaleur, diminution du métabolisme. Le sommeil paradoxal ne peut commencer que lorsque les systèmes aminergiques sont silencieux. Leur inhibition dépend à la fois du facteur libéré par la région préoptique et de l'arrêt du système bulbaire. Les mécanismes exécutifs du sommeil paradoxal peuvent alors entrer en jeu.
Organisation schématique des systèmes exécutifs du sommeil paradoxal
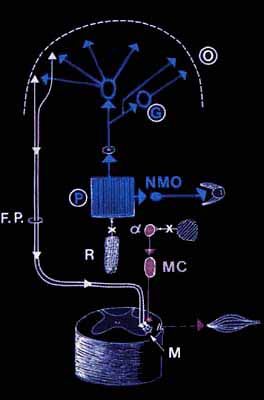
Le générateur de l'activité PGO (P) est responsable des mouvements oculaires en agissant sur les neurones oculaires (MNO). Des voies ascendantes montent vers les noyaux genouillés latéraux (G) et le cortex occipItal. (O).
C'est à ces endroits que l'on peut recueillir l'activité dite ponto-géniculo-occipItal.e (PGO), car elle présente une activité de très grande amplitude. D'autres relais conduisent l'activité du générateur jusqu'au cortex. En réponse (directe ou indirecte) à cette activité, les neurones pyramidaux (triangle) envoient des influx descendants dans le faisceau pyramidal (FP). Celui-ci devrait exciter le motoneurone médullaire (M) et déclencher un mouvement. Cela est impossible car le locus coeruleus alpha met en jeu le noyau magnocellulaire du bulbe (MC) qui par l'intermédiaire du faisceau réticulospinal va inhiber le motoneurone. Ces mécanismes ne peuvent survenir que si le système du raphé (R) et le système du locus coeruleus (LC) sont eux-mêmes inhibés.
Interaction entre système permissif et exécutif au cours du sommeil paradoxal
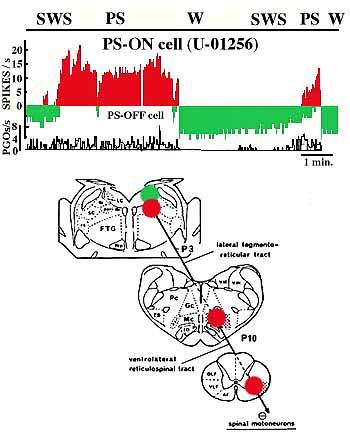
En haut, en vert, fréquence de l'activité unitaire d'un neurone du locus coeruleus (neurone PS off). L'activité de ce neurone s'arrête au début du sommeil paradoxal alors que l'activité d'un neurone du locus coeruleus alpha (en rouge) (neurone PS on), qui était totalement absente, se met à augmenter pendant la période de sommeil pour s'arrêter brusquement ensuite.
Au dessous, représentation schématique des zones d'enregistrement: en vert, le locus coeruleus, noradrénergique; en rouge, le locus coeruleus alpha (dont le neurotransmetteur est encore inconnu et la voie descendante responsable de l'inhibition du tonus musculaire avec le relais bulbaire et le faisceau réticulospinal dans la moelle
(D'après K. Sakai)