| Telencephalic and rhombencephalic sleep in the cat |
| Jouvet M. The Nature of Sleep Ciba Foundation Symposium Churchill (1961) |
A brief review of the physiological data relating to the nature of sleep enables us to get an idea of the duality of structures and mechanisms brought into play during this periodic mystery.
Structural duality : Without referring back to the opposition between cerebral and body sleep, it should be remembered simply that Von Economo (1927), basing his findings on clinical and pharmacological facts, had already contrasted the passive character of telencephalic sleep with the active regulating part played by a hypothetical mesodiencephalic centre.
Duality of mechanisms : Is it possible to reconcile, in a single mechanism, Bergson's sleep "désinterêt" with that vItal. need, the lack of which brings death? Is it possible to reconcile, without calling upon two distinct mechanisms, the hypothesis of passive sleep resulting from neuronal fatigue or deafferentation of the ascending activating reticular system (Bremer, 1954) with Pavlov's theory of sleep as a cortical active inhibition phenomenon or with Hess's (1928) hypothesis of the existence of a diencephalic sleep centre which can be brought into play through stimulation ?
Recently, the idea of sleep mechanism duality has found support from EEG analysis. Slow cortical activity was long considered as the only EEG index of sleep in the cat (Rheinberger and Jasper, 1937; Hess, Koella and Akert, 1953) but certain observations have since shown the existence of a rapid cortical activity (Rimbaud et al., 1955), the periodic recurrence of which has been compared by Dement (1958) to "activated sleep", an intermediary state between wakefulness and sleep.
A brief account will now be given of the experimental results (Jouvet, Michel and Courjon, 1959b; Jouvet, 1961) which enables us to contrast sleep accompanied by slow cerebral activity requiring the integrity of the telencephalon to sleep accompanied by rapid cortical activity, which we shall show to be dependent on the intervention of rhombencephalic structures.
Review of methods : The problem of discovering the cerebral formations responsible for the various stages of sleep can be solved only by animal experiments combining nerve destruction and stimulation. Some methods (Bremer, 1935; Batini et al., 1958) are based essentially on cortical EEG records, which are considered as the most reliable evidence of wakefulness level. A "slow" tracing becomes evidence of sleep, a "rapid" tracing of wakefulness. There are, however, many exceptions.
(1) A synchronized or "slow" tracing is not necessarily a sign of physiological sleep. For example, the irreversibility of barbiturate narcosis or coma due to reticular formation destruction enables us to eliminate one phenomenon of sleep.
(2) A "rapid" cortical tracing is not necessarily a sign of wakefulness: ether narcosis, for instance, is accompanied by "cortical activation" and there are sleep phases in the cat with rapid cortical activity.
(3) Last but not least, the study during chronic experiments of an animal whose brain stem has been sectioned between the quadrigeminal bodies results in a cortical tracing which, if considered by itself, would lead to the conclusion that the animal is asleep; in fact it reacts to pain by fleeing movements and miaowing.
The imperative necessity of checking animal sleep behaviour has lead us to record cortical and subcortical electrical activity in relation to the classic somato-vegetative manifestations of sleep (muscular tonic activity, heart beat, respiratory rhythms, etc.). In a brain stem section, the electrical activity of that part of the encephalon which is situatedbehind the lesion (and not only in front of it), and which is therefore linked to the exterior milieu by its main afferent and efferent pathways, was recorded during the periodic behavioural variations of wakefulness. It has been known since the time of Goltz (1892) that a decorticate animal can sleep. The works of Bard and Macht (1958) and Rioch (1954) have also shown that mesencephalic animals showed behavioural altemations of wakefulness and sleep.
Materials and methods
Results were obtained from 52 chronic cats ( 12 normal cats, 11 totally or partially decorticate, 20 with total or partial brain stem section, 7 with partial coagulation of the brain stem, and 2 which had undergone total ablation of the cerebellum). These animals were implanted with nickel-chrome multipolar sub cortical electrodes and steel cortical electrodes, and were all studied for more than one week. Decortication was carried out in one operation by suction after removal of the skull. An acrylic resin plastic roof was then inserted in place of the bone and the subcortical electrodes were thus kept at that level (Jouvet, Michel and Courjon, 1959a). The brain stem was sectioned by means of a stereotaxically oriented cutting blade, and coagulation was obtained by means of anodal electrolytic lesion. In some cases, the entire part of the encephalon situated in front of the midbrain was removed by aspiration. Poikilotherm, mesencephalic or pontile animals were kept warm and always studied at normal temperatures between 37° and 39°. The cats were placed in soundproof cages and EEG recording was carried out daily for 6 to 12 hours. As a routine measure, together with the EEG, the EMG was recorded by means of a bipolar electrode permanently mounted at the level of the neck muscles. The eye movements were also recorded by electrodes fixed on both sides of the eye balls. The behaviour of the animals (pupils, nictitating membranes, posture, etc.) could also be observed through glass. In all cases, the siting of the electrodes and the topography of the lesions were checked histologically.
Results
A. Intact sleeping cats (Fig.1)
Two different stages can be distinguished on the records:
(1) Falling asleep and " slow " sleep. This stage is marked by the appearance of spindles followed by slow waves which invade the cortex, the diencephalon and then the mesencephalic reticular formation (R.F.). During this stage, the animal prepares for sleep. It bends its head while the EMG of the neck muscles discreetly falls. Breathing is regular; so is the heart beat which slows down in comparison with that in the waking state. As the invasion of reticular formation by the slow waves proceeds, the threshold of arousal by direct excitation of the R.F. increases from 20 percent to 50 per cent.
(2) "Rapid" sleep phase or "paradoxical phase" (p.p.) (Jouvet, Michel and Courjon, 1959C) (Figs. 1 - 3). This always follows a " slow" sleep phase and never appears immediately after wakeful ness. It starts suddenly and is distinguished by a rapid and low voltage corticomesodiencephalic activity, identical to that of wakefulness, while the dorsal rhinencephalic formations show a slow rhythmic activity, identical to that which has been described during arousal at this level (Green and Arduini, 1953). At the same time, a 6 to 8 per second spindle activity appears at the level of the pontile R.F. The auditory cortical, and particularly reticular evoked responses undergo a marked reduction in amplitude compared with that in the waking state or during "slow sleep." This phase, 10 to 15 minutes in duration, is accompanied immediately and constantly by total disappearance of EMG activity in the recorded muscles. It is periodically repeated during behavioural sleep, with intervals of 20 to 30 minutes (Fig. 2).
During that phase, the posture of the animal is the same as that found in profound sleep. Postural muscles completely lack tone, nictitating membranes almost entirely cover the pupils which are myotic, while the eyeballs are frequently shaken by short rapid jerks. Movements of the vibrissae and, more rarely, brief jerks of the jaws and the tail can also be observed. Cardiorespiratory variations are constantly observed: breathing becomes irregular, more superficial and quicker than during the slow wave phase, while the heart beat is slowed down, or, more rarely, quickened (Fig. 3).
That this phase of sleep is more profound than the slow wave ase is supported by the following arguments :
(I) An auditory stimulation, insufficient to produce arousal, causes the spindle and the slow wave stage to reappear.
(II) The arousal threshold, measured in decibels for an auditory stimulation, is higher during this phase than during the "slow" phase.
(III) Last but not least, the behavioural threshold of arousal through direct stimulation of the mesencephalic R.F. in creases by 200 to 300 per cent compared with that of the slow stage of sleep (Jouvet, Michel and Courjon, 1959C; Benoit and Bloch, 1960).
Identical EEG and behavioural aspects during these two phases of sleep are found in partially decorticate animals or totally cerebellectomized cats (Jouvet and Michel, 1960a). In these animals, the alpha-type rigidity which distinguishes the post-operative period is abolished during p.p.
B. Cats which have undergone total removal of the neocortex (Fig. 4)
A surprising alteration in the EEG recorded at the level of the subcortical structures is produced by this operation (Jouvet and Michel, 1958). There is, in effect, a permanent lack of spindles and slow waves during the survival period of the animal (up to three months). Mesodiencephalic structures continuously exhibit a fast low voltage activity. On the other hand, the p.p. is shown by phenomena identical with those of the intact animal: appearance of spindles (6/sec.) at the level of the pontile R.F., rapid rhinencephalic activity, disappearance of EMG activity, cardiorespiratory alterations, and appearance of eye movements (Fig. 3).C. Chronic pontile or mesencephalic animal (Figs. 5 and 6)
The electrical activity of the cortical and diencephalic formations situated in front of the section continually exhibit the classic appearance described at the level of the cerveau isolé (Bremer, 1935), i.e. a continuous mixing of spindles and slow waves, whatever the state of wakefulness.
Behind the section, however, mesencephalic activity constantly remains rapid during waking. The periods of behavioural sleep are marked by the appearance of spindles at the level of the pontile R.F. and a complete disappearance of EMG activity in the neck, which is remarkable since muscular activity is greatly increased during wakefulness (decerebration hypertony) (Fig. 2) (Jouvet and Michel, 1959). During these periods, the eye movements are slower and less frequent than in decorticate or normal animals, while the nictitating membrane is relaxed. Finally, important cardiorespiratory variations are observed (Fig. 3).
The duration of these phases is similar to that observed in intact animals, i.e. 10 to 15 minutes. The interval between phases is longer : 40 to 60 minutes (Fig. 2). The behavioural arousal threshold produced by reticular stimulation (shown by an increase in hypertony and mydriasis) is also increased by 200 per cent during p.p., compared with the state of wakefulness.
D. Posterior pontile animal
In animals sectioned along a plane inclined at 60° in relation to the frontal plane, at the posterior part of the pons (between the pons and the trapezoid bodies), no periodic variation of muscular tone, respiration or heart rhythm, such as appears in mesencephalic or pontile animals during p.p., can be observed. EMG activity consistently persists during the survival of the animal (up to ten days). In front of the section cortical activity is often rapid and resembles that described by Batini and co-workers (1958) in cat sectioned in the middle of the pons. Periods of slow waves and spindles can, however, be observed for approximately 40 per cent of the time.
Topography of the systems responsible for the two stages of sleep
(1) First phase of sleep (spindles and slow waves)
Whatever the subcortical structures may be which are responsible for the synchronization phenomena (diffuse thalamic system, Jasper, 1949; or caudal region of the brain stem, Batini et al., 1958) one fact appears to be certain: the spindles and the slow waves which are observed during the first phase of sleep require the presence of the neocortex, since there are no sub cortical spindles or slow waves in the decorticate animal or from behind a section of the brain stem. The study of animals whose brain stem has been partially sectioned enables us to state that the corticifugal inhibitory descending tracts are diffuse within the brain stem R.F., because the slow waves appear behind a section, leaving part of the R.F. intact, however small.
(2) Determination of the cerebral structures responsible for the rapid activity phase (p.p)
a) Structures responsible for the triggering of this phase. Pontile EEG activity and somato-vegetative phenomena which are characteristic of the p.p. continue to appear normally in chronic decorticate, mesencephalic or pontile animals. These phenomena, however, are completely lacking in cats whose brain stems have been sectioned between the pons and the trapezoid bodies. The structures which are responsible for the triggering of such periodic phenomena must, therefore, necessarily be found at the level of the pons (Fig. 7).
The formal proof of this is provided by coagulation experiments limited to this level. These were carried out over a width of 3 to 4 mm. at the level of the pons tegmentum. The nudeus pontis caudalis, the posterior part of the nucleus pontis oralis and the superior central nudeus of Bekhterev were destroyed (Fig. 8). Not one of these animals exhibited a p.p. during sleep.
They all showed, however, a slow cortical activity during the first phase of sleep and were capable of a rapid activity during spontaneous or provoked arousal. The reticular formation of the pons thus appears to be responsible for the rapid activity phase of sleep, which can therefore be described as the rhombencephalic phase of sleep.
(b) Cerebral tracts responslble for rapid cortical activity. These tracts are certainly different from the ascending activating reticular system which is responsible for cortical "arousal" (Moruzzi and Magoun, 1949). Indeed cats whose R.F. is interrupted at the level of the midbrain tegmentum, and which are thus incapable of any rapid cortical activity during nociceptive stimulation or reticular stimulation behind the lesions, continue, however, to exhibit rapid cortical activity during p.p. if the ventral part of the mesencephalon is not damaged at the level of the interpeduncular nucleus and the mammillary bodies (Jouvet and Michel, 1960a). Such structures are known to be passages in a circuit (limbic midbrain circuit-Nauta, 1958) linking certain nuclei situated at the level of the pontile R.F. to limbic and rhinencephalic formations.
It is thus probable that such a circuit is responsible for some cortical electrical aspects of the p.p., particularly for the rhinencephalic rhythmic activity.
c) Structures responsible for somato-vegetative phenomena. The appearance of typical p.p.'s in the cerebellectomized animal having disposed of any possibility of the cerebellum playing a part, the somato-vegetative phenomena associated with this phase may be explained by the active intervention of the bulbar inhibitory R.F. (Magoun, 1950), which is known to exert a general control on muscular tone and acts on the alpha-type as well as on the gamma-type rigidities (decerebellation and decerebration rigidities). Respiratory and cardiovascular centres are also known to exist at the level of the pons and their intervention would explain the characteristic variations in respiratory and cardiac rhythms.
Mechanisms of the rhombencephalic phase of sleep
This is an active phenomenon as it is possible to trigger off the appearance of the rhombencephalic phase of sleep by stimulating the pontile R.F. of the intact animal, provided that this stimulation occurs during the slow wave phase of sleep (Fig. 9) (Jouvet and Michel, 1960b).
In mesencephalic cats, stimulation of this same zone is also capable of triggering periods of "rhombencephalic sleep". Phases of 15 minutes duration have thus been obtained after stimulations lasting 1 to 2 seconds. After the animal has spontaneously awakened, a refractory period is observed, during which an identical stimulus determines a hypertonic phase accompanied by agitation. Therefore it is never possible to trigger several periods of rhombencephalic sleep successively. An interval of 15 to 20 minutes must be allowed to elapse before a new stimulation can produce a new phase of sleep. In some animals, sleep has been obtained by summation of stimuli of long duration (10 to 20 seconds) and in some cases a latent period of 30 to 60 seconds was observed between the end of the stimulation and the beginning of sleep. Controls have shown that these phenomena are not accompanied by significant variations in blood pressure.
Chlorpromazine (3 to 5 mg./kg.) and atropine (0-2 to o-3 mg./kg.) considerably reduce the duration of the p.p.'s or even stop them from appearing, particularly in mesencephalic animals. On the other hand, the injection of cholinergic drugs (eserine) produces longer p.p.'s although their frequency is not increased (Jouvet and Michel, 1960a).
These facts are difficult to explain as a whole through purely neuronal mechanisms. They lead us to submit the hypothesis of the existence of a neurohumoral mechanism which would "discharge" periodically during behavioural sleep but which could not be brought into play until a sufficient "stock" of neurohormones was gathered. The inhibiting action of such a mechanism on the awakening system (which is expressed by a very important increase in the threshold of reticular arousal) must be explained by a direct action of the " triggering rhombencephalic zone" on the ascending activating system since the awakening threshold rises in the intact as well as in the mesencephalic animal, which eliminates the intervention of possible cortico-reticular feedbacks. Although the periodic physiological mise en jeu of the rhombencephalic phase of sleep is not yet understood, its psycho-physiological significance seems important. Dement and Kleitman (1957) have shown that in man the oneiric activity occurred periodically during sleep and that it was accompanied by eye movements and by cortical low voltage electrical activity. These findings have been confirmed by us (Jouvet, Michel and Mounier, 1960). We have put forward other arguments which enable us to accept that oneiric activity occurs in man during the rhombencephalic phase of sleep. It is thus surprising to find that such important and mysterious psychic phenomena as dreams are dependent on the intervention of structures which are hidden in the lower part of the brain stem.
Conclusions
These results enable us to dissociate the two phases of sleep which are relatively independent of one another. The EEG tracing of decorticate or mesencephalic cats shows no slow phase whereas periods of "rhombencephalic sleep" persist. In the cats with a pontile R.F. lesion, on the other hand, only the phase of "slow sleep" appears, without any p.p. These facts lead us to accept the intervention of two different systems during physiological sleep in the cat.
(1) The first system appears to intervene during " spindles and slow waves" sleep in the intact animal. It requires the presence of the neocortex which is responsible for the slow activity. This corticifugal slow activity expresses an inhibition phenomenon since the arousal threshold produced by direct stimulation of the reticular formation rises during this stage. This phase may there fore be described as telencephalic sleep. It is possible that this telencephalic stage may represent a stage which is acquired during telencephalization and that it could be described as "neosleep".
(2) The phase of "rapid" sleep- (paradoxical phase) which we propose to call the "rhombencephalic phase" is dependent upon a totally different system, situated at the level of the pontile reticular formation. It controls, through the inhibitory R.F., the somato-vegetative phenomena (disappearance of all muscular tonic activity even in the cases of decerebration or decerebellation hypertony, variation in respiratory and cardiac rhythms). The rapid cortical activity which accompanies this phase is not suppressed by the interruption of the activating reticular system. This phase of sleep which only exists in mesencephalic or pontile cats could be likened to an "archisleep". This phase is more profound than the first one as the threshold of awakening is increased in comparison with that of the slow wave phase of sleep. Finally, it can be triggered off in animals by stimulating the lower part of the brain stem, and it is suggested that this phase depends upon a neurohumoral mechanism.
Discussion
Moruzzi : I should like to report some experiments which have been performed in Genoa by G. F. Rossi and his colleagues, T. Hara, E. Favale and G. Sacco. These experiments also concern the stage of sleep which is characterized, in the cat, by a flat EEG record. Hara, Favale, Rossi and Sacco (1960. Boll. Soc. Ital. Biol. sper., 36, 1203) have fully confirmed the observation by Jouvet, Michel and Courjon that the electromyogram of neck muscles is silent during this stage of sleep. They have also confirmed that the reticular responses to clicks are greatly decreased and even abolished, and that the threshold for behavioural arousal elicited by reticular stimulation is much higher. Hence they agree with the view that the sleep is deeper in these stages than when high voltage slow waves are present on the EEG record. The following observations, however, point out that there are also striking electrophysiological similarities with the conditions occurring during wakefulness: (1) the recruiting responses are abolished or strongly decreased; (2) the EEG synchronization elicited by low-rate stimulation of the brain stem reticular formation is also abolished; finally (3) an intravenous injection of thiopentone sodium (10-20 mg./kg.) will synchronize the EEG, while the electromyogram of neck muscles remains silent and the threshold for behavioural arousal is further increased. The same EEG synchronization occurs when barbiturates are injected on the background of low voltage fast activity that characterizes the pretrigeminal midpontine preparation (Batini, C., Moruzzi, G., Palestini, M., Rossi, G. F., and Zanchetti, A. [1959]. Arch. Ital. Biol., 97, 1). Dr. Jouvet, can you suggest an explanation for the effect of thiopentone sodium during the deep stage of sleep characterized by a flat EEG record ?
Jouvet : Sometimes during the paradoxical phase if you make a noise the cat will not wake up but will get to the slow wave stage of sleep (i.e. it will go back to a lighter stage of sleep).
Moruzzi : The doses of thiopentone sodium used by Rossi and his colleagues would produce typical spindles, when injected in an awake cat. When injected during deep sleep they still have a depressant influence, as shown by behavioural tests. Moreover the EMG of the neck muscles remains silent. It is puzzling that the EEG reverts to patterns that are regarded as typical of lighter stages of sleep following injection of thiopentone sodium.
Jouvet : But this is narcosis and not sleep
Bradley : You mentioned using atropine and eserine, Dr. Jouvet. It would be interesting to know what their effects are. I would suspect that they modify the electrical record without altering the behavioural pattern.
Jouvet : Yes, under atropine there is no fast activity during sleep and also there is no total disappearance of EMG activity, so that it looks as though atropine would block this mechanism, and that this mechanism is somewhat cholinergic in nature. With eserine the duration of the paradoxical stage of sleep is much longer but the periodicity remains the same. There was no behavioural change in sleep with atropine but with eserine the eye movements were very much more enhanced.
Bremer : I have very great appreciation of your work, Dr. Jouvet, and the only reservation I should like to make is: are we compelled to admit that this flat desynchronized tracing necessarily means a great activation of the cortex ? Could we not assume that the tracing is flat for other reasons but that it is not the same flattening as in arousal ? In the figure published by W. Dement (I958. Electroenceph. Clin. Neurophysiol., 10, 291) there is a distinct difference between the awakening tracing and the "activated sleep" one. Therefore it would be a relief if we could decide that the so-called paradoxical or activated sleep is not activation of the cortex, which would be a really paradoxical affair.
Jouvet : From the EEG record it is not possible to decide whether the cat is awake or in deep sleep. You are obliged to look at the EMG activity.
Figure 1
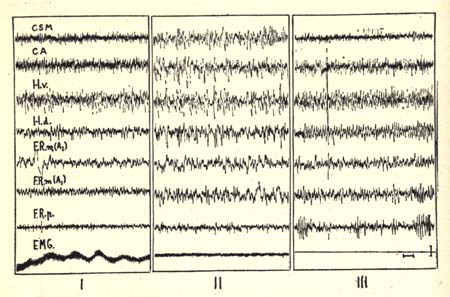
I- Arousal.
II- Slow phase of sleep: slow waves and spindles at the cortical and reticular level. Persistence of EMG activity of the neck muscles (EMG).
III- Rhombencephalic phase of sleep (paradoxical phase). Fast cortical and reticular activity. Rhythmic activity at the dorsal hippocampal level (H.d.), spindles at the pontine reticular level (F.R.p.) and total disappearance of the EMG activity.
C.S.M.; C.A.: sensory motor cortex; acoustic cortex.
H.v.: ventral hippocampus.
F.R.m.: mesencephalic medial reticular formation in frontal planes of Horsley-Clark: Al and A2.
Time scale: 1 second, 50 microvolts.
In all figures, recordings are bipolar.
Figure 2
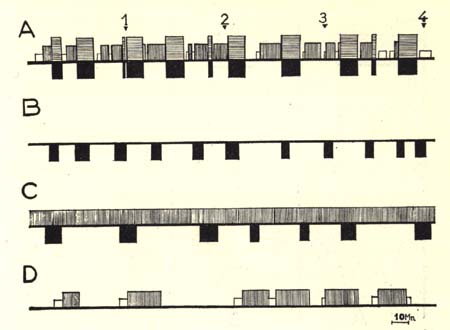
Diagrammatic representation of the sleep-wakefulness rhythm in
- (A) intact,
- (B) decorticate,
- (C) mesencephalic cats, and
- (D) cats with medial coagulation of the pontine reticular formation.
Four hours continuous recording, one week after the intervention, for each cat.
(Time scale: 10 minutes)
In black, the rhombencephalic stage of sleep with fast cortical EEG pattern in normal cats (horizontal hatching) and spindling activity at the pontine level, total disappearance of EMG activity (movements of eyes, etc.) in normal, decorticate and mesencephalic cats.
In white, spindling activity on the cortex.
Vertical hatching, slow wave and spindles at the cortical and diencephalic level.
Horizontal line, arousal.
Note the absence of the slow wave and spindle phase of sleep in de corticate cat, and the absence of the rhombencephalic phase in cat with pontine lesion. Note also the relative wakefulness of the latter type of preparation.
Figure 3
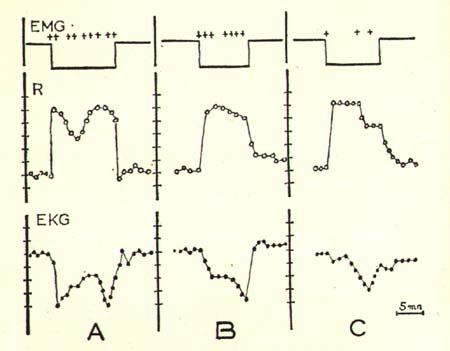
Changes of cardiorespiratory activity during the rhombencephalic phase of sleep in a normal (A), decorticate (B) and mesencephalic (C) chronic cat.
Disappearance of EMG is schematically indicated on the upper line and the crosses signal the movements of eyes.
Scale of respiration (R) = 1 per min. (for EKG = 5 min.)
Time scale: 5 minutes.
Figure 4
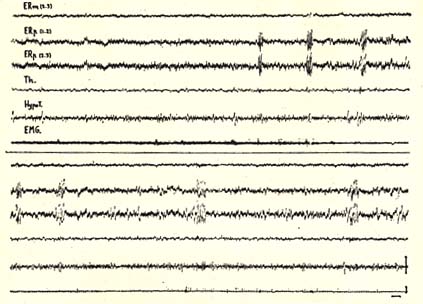
Rhombencephalic phase of sleep in a decorticate cat (X1)
The two recordings are continuous.
Appearance of a spindle-like activity in the nucleus pontis caudalis (F.Rp) and concomitant disappearance of the EMG activity of neck muscles (EMG). Note the absence of slow waves m the subcortical structures before the rhombencephalic sleep. Numbers (1-2), (2-3) refer to bipolar recording through inferior (1), middle (2) and superior (3) leads of a tripolar electrode.
F.Rm: mesencephalic reticular formation.
Th: thalamus median.
HypoT: lateral hypothalamus.
Scale: 1 sec., 50 microvolts.
Figure 5
Dorsal (A) and ventral (B) views of truncated brain stem of cat X2.
Figure 6
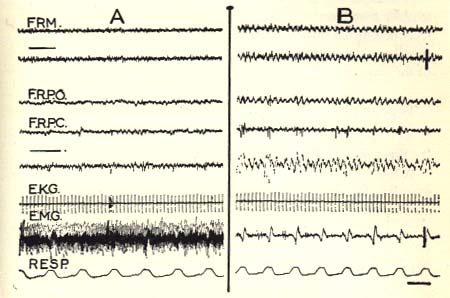
Arousal (A) and rhombencephalic sleep (B) in cat X2.
Note the rhythmic activity in the subcortical activity and the spindle-like activity in the nucleus pontis caudalis (F.R.P.C.), the slowing of E.K.G., acceleration of respiration (R) and the total disappearance of the E.M.G. activity of the neck muscles.
FRM.=Posterior mesencephalic reticular formation.
F.R.P.O.=Nucleus pontis oralis.
Figure 7
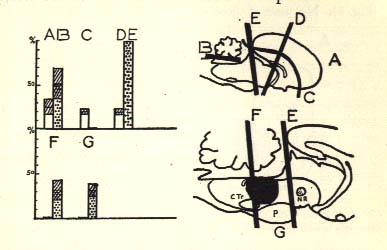
Diagrammatic representation of the sleep-wakefulness rhythm in (A) intact, (B) cerebellectomized, (D) mesencephalic, (E) pontine, (F) retropontine cats, and (G) in cats with limited medial lesion of the pontine reticular formation (in black) (cf. Fig. 8).
Numbers at left indicate the percentage, for 12 hours continuous recording, of either synchronized and slow activity (columns with dots) or fast activity and disappearance of EMG activity during sleep (white columns). Bach category represents the mean of sixteen recordings in four cats. Cross-hatching indicates standard deviation. Note the absence of synchronized activity in decorticate cat (C), and the absence of the rhombencephalic stage of sleep in F and G. In mesencephalic and pontine cats (D, E), the rhombencephalic stage of sleep does not induce fast activity at the cortical level and is distinguished on the BMG and by the pontine activity.
N.R.: Red nucleus.
P: Pons.
C. Tr.: Trapezoid body.
Figure 8
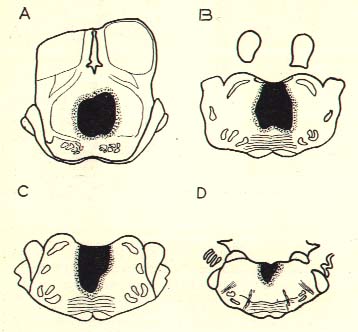
Cat C3. Examples of lesions in nucleus pontis caudalis shown at planes P. 5, 6, 7 and 8.
This cat did not show any fast activity or disappearance of EMG activity during sleep, during 10 days of survival. He had the slow phase of sleep and a normal fast activity during arousal. He could walk and feed himself
Figure 9
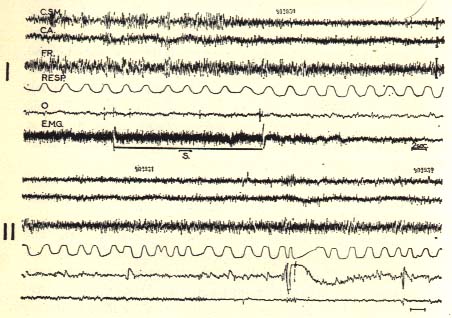
Triggering of the rhombencephahc phase of sleep in an intact chronic cat (V2)
The horizontal line (S) signals the stimulation of the pontine reticular formation (300/sec.,1 millisecond, 0-80 volt, bipolar) for 11 seconds during the slow phase of sleep.
Note the appearance of fast activity at the cortical and reticular level, the total disappearance of the EMG activity of the neck muscles (E.M.G.), the acceleration and irregularity of the respiration (RESP.), and the appearance of rapid eye movements (O). This period of sleep lasted for 10 minutes.
The same phenomenon was reproduced five times in this recording session
The two recordings I and II are continuous.
C.S.M.; C.A.: sensory motor cortex; acoustic cortex.
F.R.: mesencephalic reticular formation.
Scale: 2 seconds, 50 microvolts.
Note the utility of polygraphic recording without which the fast cortical activity might be interpreted as an "arousal".