| Phylogenèse des états de sommeil |
|
Michel Jouvet Acta Psychiat. Belg., 94, 256-267 (1994) |
Phylogeny of the Stafes of Sleep
The states of sleep in man (orthodox and paradoxical sleep) are the achievement of a long evolution which has successively integrated several mechanisms. The circadian clock (predictive homeostasis a system responsible for saving energy utilisation, a mechanism which adapts the duration of sleep to the preceding duration of waking (reactive homeostasis) and finally the apparition of an ultradian pacemaker in homeothermic mammals which will be responsiblef or dreaming in man.
Phylogenèse des états de sommeil
Le sommeil de l'homme est l'aboutissement d'une très longue évolution phylogénétique qui a intégré les mécanismes suivants:
- L'"invention" chez les êtres unicellulaires de mécanismes "d'homéostasie prédictive". Ces mécanismes sont commandés par une horloge interne, d'à peu près 24 heures (circadienne).
- Des processus d 'économie d'énergie: Inactivité, repos et sommeil, permettent en effet de réduire la consommation d'énergie et de nourriture. Dans certaines niches écologiques, un processus saisonnier, l'hibernation remplace l'alternance quotidienne du sommeil en réduisant considérablement la consommation d'énergie par diminution de la température corporelle jusqu'à 1 à 2°C.
- Des processus d'homéostasie réactive. Plus la période d'activité est Iongue ou intense, plus le sommeil "récupérateur" sera également long ou intense.
- Enfin, depuis l'apparition de l'homéothermie avec les oiseaux,
le sommeil sert également à protéger et préparer
un nouvel état survenant avec une périodicité ultradienne,
le sommeil paradoxal, (SP) ou sommeil avec mouvements rapides oculaires,
qui est chez l'homme le substratum neurobiologique de l'activité
onirique.
Nous passerons d'abord en revue l'apparition et l'évolution de ces mécanismes au cours de l'évolution phylogénétique, avant de montrer comment on en retrouve les traces au niveau du sommeil d'un sujet humain.
I. Phylogenèse des états de sommeil
A) Les êtres unicellulaires et les plantes
 En
1729, un astronome, Jean-Jacques d'Ortous de Mairan, enferme une sensitive
(mimosa pudica) dans un cabinet noir. Il remarque alors que, même
en l'absence d'alternance lumière-obscurité, les feuilles
de la sensitive continuent à se redresser ou de se replier toutes
les 24 heures. Il en conclut donc que cette plante avait une "horloge
interne" qui lui donnait l'heure.
En
1729, un astronome, Jean-Jacques d'Ortous de Mairan, enferme une sensitive
(mimosa pudica) dans un cabinet noir. Il remarque alors que, même
en l'absence d'alternance lumière-obscurité, les feuilles
de la sensitive continuent à se redresser ou de se replier toutes
les 24 heures. Il en conclut donc que cette plante avait une "horloge
interne" qui lui donnait l'heure.
Ce phénomène est universel puisqu'il existe également au niveau de l'activité de certaines enzymes chez des êtres unicellulaires (Acetabularia Euglena). Les mécanismes moléculaires de cette horloge sont encore inconnus. Ainsi, bien qu'il soit évidemment impossible de parler d'activité repos chez les algues et les plantes, il faut savoir qu'il existe dès l'apparition de la vie une horloge moléculaire circadienne chez tous les êtres vivants.
B) Les états "voisins du sommeil" chez les invertébrés
Il est bien connu que les insectes ont un rythme journalier de repos et d'activité. Ce rythme n'est pas simplement le reflet de l'alternance lumière obscurité car il peut continuer à être observé même si l'environnement ne fournit aucune indication de temps qui puisse servir de synchroniseur (ou Zeitgeber). Les preuves de l'existence d'un état proche du sommeil ont été trouvées sur des critères comportementaux et physiologiques, aussi bien chez le scorpion que chez l'abeille. Les corrélats du repos sont l'immobilité, une posture caractéristique, la diminution de la température, la baisse du tonus des muscles de la nuque, l'élévation du seuil d'éveil et une augmentation de la sensibilité des neurones visuels. Le scorpion ou la blatte passe de la position érigée, abdomen et tête redressés, à une position où le corps et la tête reposent à plat sur le sol. Chez l'abeille, la tête est inclinée et le tonus musculaire de la nuque réduit. Le seuil d'éveil, par des stimulations extérieures, s'élève. Dans le stade d'immobilité le plus profond, les antennes de l'abeille sont repliées contre la tête. Il semble qu'une régulation homéostasique commence à apparaître chez les insectes puisque des indices de mécanismes compensatoires ont été constatés aussi bien chez la blatte que le scorpion. En effet, le repos comportemental est plus long au cours de la récupération chez ces 2 espèces si la période d'activité est allongée (par des stimulations mécaniques, par exemple).
C) Les états d'activité et d'inactivité chez les vertébrés polkilothermes
(Bauchot, 1984; Karmanova, 1982; Tobler, 1992)
Les poissons:
 Dans
ce domaine, la majorité des recherches est seulement comportementale.
L'existence d'états ressemblant au sommeil a été
prouvée chez de nombreuses espèces. Certaines s'enfoncent
dans le sable pour y passer la nuit. D'autres changent de couleur la nuit.
Certains poissons perroquets peuvent secréter une enveloppe muqueuse
dans laquelle ils restent sans bouger toute la nuit. Au cours de ces états
de repos, les poissons réagissent plus lentement à des stimulations
sensorielles. Chez la tanche (Tinca Tinca), on a pu enregistrer à
la fois l'activité électrique du cerveau en même temps
que l'activité des branchies et des différents muscles,
le rythme respiratoire, le rythme cardiaque et enfin la réactivité
à des stimulations sensorielles. Il n'existe pas de différences
notables entre l'activité électrique rapide et de bas voltage
du cerveau pendant l'éveil (nocturne) ou pendant le repos (diurne).
D'autre part, à aucun moment il n'a été vu, au cours
des longues périodes d'inactivité chez la tanche, de critères
permettant de supposer qu'il puisse exister des périodes de mouvements
oculaires accompagnées de variations végétatives.
Rien ne permet donc d'afffirmer qu'un état périodique, analogue
au sommeil paradoxal, soit présent chez les poissons.
Dans
ce domaine, la majorité des recherches est seulement comportementale.
L'existence d'états ressemblant au sommeil a été
prouvée chez de nombreuses espèces. Certaines s'enfoncent
dans le sable pour y passer la nuit. D'autres changent de couleur la nuit.
Certains poissons perroquets peuvent secréter une enveloppe muqueuse
dans laquelle ils restent sans bouger toute la nuit. Au cours de ces états
de repos, les poissons réagissent plus lentement à des stimulations
sensorielles. Chez la tanche (Tinca Tinca), on a pu enregistrer à
la fois l'activité électrique du cerveau en même temps
que l'activité des branchies et des différents muscles,
le rythme respiratoire, le rythme cardiaque et enfin la réactivité
à des stimulations sensorielles. Il n'existe pas de différences
notables entre l'activité électrique rapide et de bas voltage
du cerveau pendant l'éveil (nocturne) ou pendant le repos (diurne).
D'autre part, à aucun moment il n'a été vu, au cours
des longues périodes d'inactivité chez la tanche, de critères
permettant de supposer qu'il puisse exister des périodes de mouvements
oculaires accompagnées de variations végétatives.
Rien ne permet donc d'afffirmer qu'un état périodique, analogue
au sommeil paradoxal, soit présent chez les poissons.
Les amphibiens: Les données concernant le sommeil des amphibiens sont encore rares. Certaines espèces ne manifestent pas de baisse de la vigilance pendant le repos comportemental (la grenouille-taureau), tandis que chez la grenouille arboricole, la réactivité aux stimulations sensorielles décroît au cours du repos. Comme chez les poissons, l'enregistrement de l'activité électrique cérébrale n'a pas permis de découvrir des différences significatives entre les périodes d'activité et d'inactivité, et il n'a pas été possible non plus de mettre en évidence de périodes de mouvements oculaires au cours des stades de quiescence.
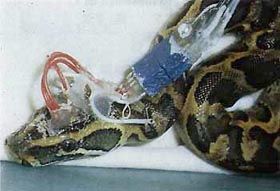 Les
reptiles: Le sommeil de certains chéloniens (tortues), crocodiliens
(alligators et caïmans), sauriens (lézards et serpents), a
été étudié avec des enregistrements polygraphiques
continus de l'électroencéphalogramme, du tonus musculaire,
des mouvements oculaires et des rythmes cardiaques et respiratoires. L'EEG
des reptiles présente quelques différences entre l'éveil
et l'inactivité. Il devient alors plus lent et surtout apparaissent
des ondes pointues de grande amplitude. Il est donc possible, chez les
reptiles, de reconnaître le sommeil par le seul examen de l'EEG
(ce qui est impossible chez les poissons et les amphibiens). Des épisodes
associant mouvements oculaires rapides et variations du rythme cardiaque,
d'une très grande brièveté (quelques secondes) et
d'une grande rareté (quelques minutes sur des enregistrements de
plusieurs semaines), ont été enregistrés chez des
crocodiliens et des lézards. L'analogie de ces épisodes
avec le sommeil paradoxal est encore très discutée.
Les
reptiles: Le sommeil de certains chéloniens (tortues), crocodiliens
(alligators et caïmans), sauriens (lézards et serpents), a
été étudié avec des enregistrements polygraphiques
continus de l'électroencéphalogramme, du tonus musculaire,
des mouvements oculaires et des rythmes cardiaques et respiratoires. L'EEG
des reptiles présente quelques différences entre l'éveil
et l'inactivité. Il devient alors plus lent et surtout apparaissent
des ondes pointues de grande amplitude. Il est donc possible, chez les
reptiles, de reconnaître le sommeil par le seul examen de l'EEG
(ce qui est impossible chez les poissons et les amphibiens). Des épisodes
associant mouvements oculaires rapides et variations du rythme cardiaque,
d'une très grande brièveté (quelques secondes) et
d'une grande rareté (quelques minutes sur des enregistrements de
plusieurs semaines), ont été enregistrés chez des
crocodiliens et des lézards. L'analogie de ces épisodes
avec le sommeil paradoxal est encore très discutée.
En conclusion, poissons, amphibiens et reptiles ne règlent pas leur température centrale, ils sont donc appelés poikilothermes ou ectothermes. Il existe de très fortes présomptions pour que l'état d'inactivité chez ces animaux soit déjà du sommeil puisque l'on a pu mettre en évidence des indices de régulation homéostatique chez des perches, des carpes et des goujons soumis à des privations de repos. L'activité forcée pendant des durées de 6 à 24 heures a ensuite été suivie d'un allongement de repos chez les 3 espèces. De même, des reptiles (caïmans, tortues, iguanes) présentent à la fois une augmentation de la durée du repos et peut-être de l'intensité du sommeil (augmentation de la fréquence des pointes au niveau de l'EEG) après des périodes d'activité prolongées artificiellement. Cependant, il n'existe aucune preuve comportementale ou polygraphique de l'existence de sommeil paradoxal chez les poikilothermes.
D) Les états de sommeil chez les vertébrés homéothermes
- Le sommeil chez les oiseaux
- Le sommeil chez les mammiferes
- Sommeil et hibernation
- Le sommeil des mammiferes marins
- Ontogenèse du sommeil et du rêve
Le sommeil chez les oiseaux
1) Le sommeil chez les oiseaux:
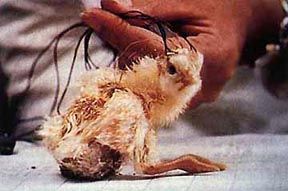 Les
oiseaux (comme les mammifères) sont des homéothermes, c'est-à-dire
qu'ils régulent leur température interne autour de 42 °C
pour la plupart des espèces.
Les
oiseaux (comme les mammifères) sont des homéothermes, c'est-à-dire
qu'ils régulent leur température interne autour de 42 °C
pour la plupart des espèces.
Une douzaine d'espèces d'oiseaux a été étudiée avec des méthodes polygraphiques. Il existe des différences EEG notables entre l'éveil (activité rapide et de bas voltage) et le sommeil (activité lente de haut voltage). Au cours du sommeil comportemental, il existe des preuves indiscutables de l'apparition périodique du sommeil paradoxal. Il se caractérise, comme chez les mammiferes, par une activation de l'EEG, par des mouvements oculaires rapides et par une diminution, parfois non totale, du tonus musculaire. Ces épisodes sont très brefs (10 à 20 secondes) mais se répètent souvent au cours de la nuit.
Il existe des différences importantes dans l'organisation du sommeil chez differentes espèces. Pendant les migrations, l'albatros ou le martinet peuvent voler sans arrêt pendant de très longues périodes. Il est alors possible que ces oiseaux ne dorment alors que d'un oeil et presentent un sommeil hémisphérique unilatéral (comme le dauphin, voir plus bas).
L'horloge biologique des oiseaux est située au niveau de la glande pineale. Elle est responsable de l'organisation circadienne du rythme activité-sommeil chez de nombreuses espèces. La régulation homéostasique du sommeil a été mise en évidence, après veille prolongée, chez des colombes et le pigeon.
En conclusion, puisque les oiseaux et les mammifères semblent avoir évolué indépendamment sans ancêtre reptilien commun, l'apparition certaine du sommeil paradoxal chez les oiseaux implique que cet état soit apparu indépendamment au cours de l'évolution.
Le sommeil chez les mammifères
2) Le sommeil chez les mammifères: Un point crucial dans l'évolution est la divergence entre les phylums des thériens, ou mammifères vivipares (marsupiaux et placentaires) et des non-thériens, ou mammiferes ovipares (monotrèmes).
a) Mammiferes ovipares (monotrèmes): chez l'echnidé et l'ornithorynque, il n'a pas été possible de mettre en évidence d'apparition périodique de sommeil paradoxal alors que le sommeil à ondes lentes est évident.
b) Mammiferes vivipares terrestres (marsupiaux et placentaires): Chez les marsupiaux (oppossum et kangourous) et 14 des 18 ordres de mammiferes placentaires étudiés par des techniques polygraphiques, le sommeil s'accompagne de variations électroencéphalographiques évidentes permettant de distinguer 2 stades à l'intérieur du sommeil "orthodoxe" (sommeil léger et profond). Chez les primates (gorilles, chimpanzés), l'organisation du sommeil orthodoxe est presque identique à celle de l'homme et comprend 4 stades.
Chez tous les ordres étudiés, le sommeil paradoxal existe. Il est très facile à reconnaître par ses critères classiques: activation de l'EEG, suppression du tonus musculaire, mouvements rapides des yeux.
Selon les espèces, il existe une grande variété dans l'organisation du sommeil par rapport à l'alternance lumière-obscurité, la durée totale du sommeil et enfin la durée de la période ultradienne du sommeil paradoxal (appelé encore T').
Chez l'homme adulte, aussi bien que chez la plupart des primates (chimpanzés, gorilles et orangs-outans), le sommeil ne comporte en général qu'une période nocturne, il est dit monophasique. Contrastant avec le sommeil monophasique des primates, celui de la plupart des autres mammiferes est polyphasique, c'est-à-dire que la période d'activité est entrecoupée d'épisodes de sommeil. Par exemple, les chats dorment beaucoup dans la journée mais ils passent également une bonne partie de la nuit à dormir. Les rats, qui sont des animaux nocturnes, dorment environ 80% des heures diurnes mais aussi 20% la nuit. On voit rarement les gros herbivores, tels que les vaches, chevaux et moutons, en train de dormir dans la journée.
c) Durée totale du sommeil (Campbell et Tobler, 1984): Il existe des différences importantes entre la durée totale du sommeil selon les espèces, même à l'intérieur d'un même ordre. Ainsi, dans l'ordre des rongeurs, le sommeil total (en % du nycthémère) occupe 50% et 65% chez le rat et le hamster et seulement 32% chez le cobaye.
De nombreuses corrélations ont été cherchées entre la durée du sommeil et certaines caractéristiques. En général, les animaux chasseurs (carnivores) ont un sommeil plus long que les chassés (herbivores).
d) Durée des cycles de sommeil (T') (Zepelin et Rechtschaffen, 1974): Cette variable est en général calculée depuis le début d'un épisode de sommeil paradoxal jusqu'au début du prochain épisode. Il existe une corrélation négative entre la durée du T' et le métabolisme alors que celui-ci est corrélé inversement avec le poids du corps ou du cerveau. Ainsi, le T' d'une souris est de 10 minutes, d'un chat de 24 minutes, d'un chien de 30 minutes, de l'homme de 90 minutes et de l'éléphant de 120 minutes.
e) Rapport cyclique (T'/DSP Jouvet, 1994): Ce rapport s'obtient en divisant la durée de la période T' par la durée moyenne des épisodes de SP. Il est voisin de 4 dans la grande majorité des espèces (sauf le lapin et les macaques). Il existerait ainsi une constante biologique selon laquelle il existerait, à l'intérieur d'un cycle de sommeil, une période "réfractaire" qui serait 3 fois plus longue que la durée du SP.
f) Régulation homéostatique des états de sommeil (Borbely, 1982): L'allongement de l'éveil (par des méthodes instrumentales ou pharmacologiques) entraîne chez tous les mammiferes une augmentation secondaire de la durée ou de l'intensité du sommeil. Il est possible également de ne supprimer que le sommeil paradoxal en gardant les animaux dans des conditions où ils ne peuvent relâcher totalement leur tonus musculaire (par exemple sur un petit support entouré d'eau, un rat ou un chat peut dormir sans présenter de sommeil paradoxal). Après une telle privation "spécifique", il existe une augmentation sélective du sommeil paradoxal alors qu'il n'y a pas de mécanisme compensateur au niveau de l'intensité des ondes lentes.
Sommeil et hibernation
3) Sommeil et hibernation (Walker et al, 1979): L'hibernation est considérée comme une adaptation physiologique qui permet aux hibernants de survivre en hiver à des périodes d'absence de nourriture, grâce à une diminution de la température centrale et donc du métabolisme. L'hibernation est un état qui se reproduit chaque année à la même saison chez des espèces de mammifères tels que les hamsters, marmottes, hérissons et certains ecureuils. Elle s'accompagne d'une baisse considérable de la température interne, jusqu'à 1°. Seuls les hibernants peuvent survivre à une telle température, alors qu'une température au-dessous de 17°C est léthale pour tous les autres mammiferes. Les animaux entrent en hibernation par augmentation du sommeil orthodoxe, tandis que le sommeil paradoxal disparaît graduellement. Quand la température centrale descend au-dessous de 25 °C, l'EEG devient plat. Les périodes d'hibernation peuvent durer plusieurs jours pendant la saison froide mais elles sont interrompues par des éveils périodiques "imprévisibles" au cours desquels la température centrale remonte jusqu'à 37 °C.
Le sommeil des mammifères marins
4) Le sommeil des mammifères marins (Mukhametov, 1984):
 L'adaptation
écologique des mammifères marins a entraîné
de très remarquables singularités de leur sommeil. Ainsi,
les cétacés ont une respiration volontaire (un dauphin anesthésié
meurt). L'évolution a donc dû choisir entre le dilemme de
rester éveillé pour respirer ou de mourir en dormant. Chez
la baleine globicéphale, certains dauphins d'eau douce ou de milieu
marin et chez un sirénien (dugong), le sommeil est unilatéral.
L'EEG d'un hémisphère présente des ondes lentes alors
que l'oeil controlatéral est fermé, tandis que l'EEG de
l'autre hémisphère présente une activité rapide
caractéristique (et que l'oeil controlatéral est ouvert).
En général, un épisode de sommeil unilatéral
dure 20 à 30 minutes et vice versa. Ces animaux peuvent ainsi contrôler
leur respiration avec l'hémisphère éveillé.
Bien qu'ils présentent des signes de sommeil unilatéraux
évidents au point de vue EEG, les dauphins peuvent continuer à
nager et ils n'arrêtent jamais leurs mouvements. Il n'a pas été
possible de prouver l'existence de sommeil paradoxal chez les dauphins.
Cependant, on peut se demander si des périodes de sommeil paradoxal
unilatérales ne pourraient pas coexister avec un éveil controlatéral
car les autres signes spécifiques du sommeil paradoxal pourraient
ne pas apparaître chez ces animaux qui n'ont pas de mouvements oculaires.
L'absence possible de sommeil paradoxal chez les cétacé
représente une des énigmes les plus importantes de la phylogenèse
du sommeil. Il ne semble pas que cela soit dû à la niche
écologique où vit le dauphin puisque le sommeil paradoxal
peut être présent chez certains phoques quand ils dorment,
non seulement à terre mais aussi dans l'eau.
L'adaptation
écologique des mammifères marins a entraîné
de très remarquables singularités de leur sommeil. Ainsi,
les cétacés ont une respiration volontaire (un dauphin anesthésié
meurt). L'évolution a donc dû choisir entre le dilemme de
rester éveillé pour respirer ou de mourir en dormant. Chez
la baleine globicéphale, certains dauphins d'eau douce ou de milieu
marin et chez un sirénien (dugong), le sommeil est unilatéral.
L'EEG d'un hémisphère présente des ondes lentes alors
que l'oeil controlatéral est fermé, tandis que l'EEG de
l'autre hémisphère présente une activité rapide
caractéristique (et que l'oeil controlatéral est ouvert).
En général, un épisode de sommeil unilatéral
dure 20 à 30 minutes et vice versa. Ces animaux peuvent ainsi contrôler
leur respiration avec l'hémisphère éveillé.
Bien qu'ils présentent des signes de sommeil unilatéraux
évidents au point de vue EEG, les dauphins peuvent continuer à
nager et ils n'arrêtent jamais leurs mouvements. Il n'a pas été
possible de prouver l'existence de sommeil paradoxal chez les dauphins.
Cependant, on peut se demander si des périodes de sommeil paradoxal
unilatérales ne pourraient pas coexister avec un éveil controlatéral
car les autres signes spécifiques du sommeil paradoxal pourraient
ne pas apparaître chez ces animaux qui n'ont pas de mouvements oculaires.
L'absence possible de sommeil paradoxal chez les cétacé
représente une des énigmes les plus importantes de la phylogenèse
du sommeil. Il ne semble pas que cela soit dû à la niche
écologique où vit le dauphin puisque le sommeil paradoxal
peut être présent chez certains phoques quand ils dorment,
non seulement à terre mais aussi dans l'eau.
Ontogenèse du sommeil et du rêve
5) Ontogenèse du sommeil et du rêve (Jouvet-Mounier et al., 1970): L'évolution pré- ou post-natale des états de sommeil a seulement été étudiée par des méthodes polygraphiques chez les oiseaux et les mammi feres. Plus un mammifere est immature à la naissance (nidicole) (chaton raton), plus il présente un état particulier, appelé "sommeil sismique". Cet état est caractérisé par des petites secousses des muscles du corps, des mouvements des yeux et une certaine hypotonie. Il existe également un "sommeil calme" et enfin de courtes périodes d'éveil, au cours desquelles le chaton ou le raton peut s'approcher de sa mère et téter. Pendant la première semaine post-natale, il n'existe pas de modification de l'EEG permettant de dissocier l'éveil des 2 états de sommeil. A partir du 8ème jour, il devient plus facile de caractériser à la fois le sommeil paradoxal (qui prend la place du sommeil sismique) et le sommeil orthodoxe qui prend la place du sommeil calme. Chez les espèces dites nidifuges, comme le cobaye, il est facile de reconnaître les 2 états de sommeil, grâce à l'EEG, dès la naissance car le cerveau est presque achevé. Par contre, un foetus de cobaye, présente égale ment un sommeil sismique. Chez le poussin qui vient d'éclore, on constate également une augmentation très nette du sommeil paradoxal.
Le sommeil paradoxal du nourrisson humain est très important après la naissance puisqu'il peut constituer jusqu'à 60% du sommeil comporte mental. Il s'accompagne, quelques heures après la naissance, de mimiques de sourire, de tristesse ou de dégoût, alors qu'un nouveau-né humain ne sourit à sa mère que quelques semaines plus tard.
Enfin, même chez les espèces monophasiques (comme l'homme), le sommeil est polyphasique au cours des premières années.
En résumé, l'ontogenèse ne semble pas récapituler la phylogenèse des états de sommeil. Il semble en effet que le sommeil paradoxal soit très augmenté quelques jours après la naissance pour décroître à des valeurs adultes en 3 semaines chez le chaton et en 4 à 5 ans chez l'homme.
La nature du sommeil sismique, qui existe au cours des derniers jours de l'organisation du cerveau, fait encore l'objet de discussion. Pour certains, il s'agirait déjà du sommeil paradoxal. Pour d'autres, il s'agit d'un état particulier accompagnant la fin de la maturation (et de la neurogenèse cérébrale). Il ne semble pas, en tout cas, exister encore de régulation homéostasique du "sommeil sismique", qui est impossible à supprimer par des drogues ou des lesions qui suppriment le sommeil paradoxal quelques jours plus tard chez le chaton (Adrien, 1976).
II. Une nuit de sommeil chez un homme de 20 ans
Selon l'aspect du tracé EEG, et de l'activité des muscles, on peut reconnaître la succession des différents stades ou états suivants au cours du sommeil comportemental. L'endormissement (stade I) s'accompagne d'une disparition du rythme alpha de l'éveil et d'une activation de l'électroencéphalogramme. Dès ce stade, la conscience est abolie et nul ne peut apprendre au cours du sommeil. Ce stade ne dure que quelques minutes. Progressivement, au cours des 90 premières minutes, des stades de plus en plus profonds de sommeil vont apparaître: le stade II, qui s'accompagne de grandes ondes phasiques, appelées K complexes, suivi des stades III et IV (appelés encore sommeil profond). Le stade II se caractérise aussi par l'apparition d'ondes rythmiques, appelées "fuseaux", tandis que les stades III et IV sont occupés par l'apparition d'ondes lentes de haut voltage, d'une fréquence comprise entre 0,5 et 3 hertz. Cette période de sommeil s'accompagne d'une persistance du tonus musculaire, de l'absence de mouvements oculaires et d'érection, d'une diminution des fréquences cardiaque et respiratoire, d'une diminution de la température centrale aux dépens d'une élévation de la température cutanée par vasodilatation.
Vers la 90ème minute, apparaît un nouvel état appelé sommeil paradoxal. Il se caractérise par l'activation de l'électroencéphalogramme, comparable au stade de l'endormissement (stade I), par une disparition totale du tonus musculaire, par des irrégularités respiratoires et cardiaque et surtout par des mouvements oculaires rapides, latéraux ou verticaux. La peau devient froide. Enfin, s'installe une érection (*) Pendant cet état, le seuil d'éveil est au moins aussi élevé que pendant le stade de sommeil le plus profond qui le précède (stade IV). La période de sommeil paradoxal dure environ 20 minutes et se termine souvent par un bref mouvement du corps tandis que l'érection disparaît rapidement. Ainsi se termine le premier cycle de sommeil.
Le sommeil "orthodoxe" survient à nouveau (pendant 90 minutes) avec réapparition des stades III et IV. La durée du deuxième épisode de sommeil paradoxal est de 25 minutes. En général, le sommeil lent profond III et IV ne réapparaît plus ensuite au cours des 3ème et 4ème cycles de sommeil.
Ainsi, une nuit de sommeil de 8 heures s'accompagne de 4 à 5 périodes de sommeil paradoxal, d'une vingtaine de minutes environ soit 100 minutes. Le sommeil profond du début de la nuit dure également à peu près 100 minutes.
On sait maintenant que le sommeil paradoxal est un état différent du "sommeil orthodoxe" qui le précède. Cet état correspond à l'activité onirique. Sa périodicité ultradienne (90 minutes) est remarquablement stable. Les exemples suivants vont nous permettre de retrouver chez l'homme les autres mécanismes acquis au cours de l'évolution.
* Pendant longtemps on a pensé que l'érection pénienne au cours du sommeil paradoxal n'existait que chez l'homme. En fait, el]e vient d'être également démontrée chez le rat. Les mécanismes et la fonction de l'érection au cours du sommeil demeurent inconnus (Schmidt et al, 1994).
A) L'horloge circadienne endogène
Nous allons nous coucher lorsqu'il fait nuit mais ce n'est pas l'alternance jour-nuit qui règle notre sommeil nocturne. Des expériences d'isolement "hors du temps", dans des grottes ou des bunkers, ont révélé qu'un sujet humain, sans repère temporel, s'endort chaque jour à peu près à la même heure. Il existe donc une horloge interne circadienne qui est réglée à peu près à 24,5 heures et qui est responsable du rythme veille-sommeil. Cette horloge, chez tous les mammifères, est située au niveau des noyaux suprachiasmatiques et reçoit des informations "photiques" de la rétine. Normalement, la vie sociale et l'alternance jour-nuit n'est qu'un synchroniseur de cette horloge ou un "donneur de temps" (Zeitgeber). La désynchronisation entre le temps local et celui de l'horloge interne, qui apparaît au cours des voyages intercontinentaux, entraîne l'apparition du sommeil au cours de la journée (JET LAG). Chez l'homme, l'horloge est responsable de processus "d'homéostasie prédictive". Comme sur les appareils extrêmement complexes, il faut que de nombreux systèmes soient mis en marche progressivement avant d'être efficaces. Ainsi, la sécrétion de cortisol est déclenchée vers 3 heures du matin par l'horloge interne pour atteindre un niveau maximum lors de l'éveil et permettre ainsi à l'organisme de faire face à toutes les demandes accrues d'énergie (Moore-Ede, 1986).
B) Economie d'énergie
Les premiers cycles de sommeil s'accompagnent d'une diminution de la température et de la consommation de glucose et d'oxygène par le cerveau. Le métabolisme général de l'organisme est plus bas que pendant le repos au cours de l'eveil. En outre, le début du sommeil s'accompagne de libération d'hormones de croissance qui jouent un rôle anabolique important.
C) Régulation homéostatique du sommeil
Chacun de nous sait bien que s'il n'a pas dormi pendant 24 heures, il dormira ensuite à la fois plus longuement et plus profondément. Ce facteur intensité peut être mesuré à la fois par des analyses de fréquence (Transformé de Fourier) et de la "puissance" des ondes lentes du sommeil "récupérateur". Dans certains cas, cette régulation homéostatique va se faire aussi bien aux dépens de l'intensité du sommeil à ondes lentes que de la durée du sommeil paradoxal. L'augmentation compensatrice de celui-ci, qui est appelée "rebond", se fait aux dépens de la durée des épisodes sans altérer la période de 90 minutes chez l'homme (Borbély, 1982).
III. Conclusion
Mis à part les cétacés et les monotrèmes, on peut admettre que la totalité des mammifères présente un sommeil comportemental qui comporte 2 états: le sommeil à ondes lentes, ou sommeil orthodoxe, et le sommeil paradoxal. Chez la plupart des mammifères, il existe également une régulation homéostasique du sommeil. Elle peut prendre différents aspects, soit par l'intensité de l'activité lente au début de la récupération, soit par l'augmentation de la durée du sommeil paradoxal, soit aux dépens des 2 états de sommeil.
Pourquoi le sommeil paradoxal apparaît-il chez les oiseaux et les euthériens, sans doute à partir de l'endothermie ? L'endothermie, qui permet une plus grande liberté par rapport à l'environnement, s'accompagne de la perte d'une faculté particulière, que possède les ectothermes adultes, c'est-à-dire a persistance de la division des cellules nerveuses ou neurogenèse. Une neurogenèse persiste chez les espèces poïkilothermes adultes (poissons, amphibiens et reptiles). Elle pourrait ainsi assurer la programmation génétique des différences inter-individuelles (phénotypiques) des comportements. Cette fonction de programmation s'effectuerait également, par division des neurones, chez les mammifères nouveau-nés au cours du sommeil sismique. A partir du moment où les neurones cessent de se diviser, quelques jours après la naissance chez les mammiferes, il faut supposer qu'un nouveau système de programmation génétique apparaisse. Cette fonction serait alors assurée par le sommeil paradoxal, qui jouerait le rôle de programmation génétique itérative de l'individualité psychologique (Jouvet, 1991).
L'arbre phylogénique du sommeil et du rêve est encore bien incomplet. Il faut espérer que le développement des techniques d'enregistrement par télémétrie, "sur le terrain", puissent permettre d'obtenir plus de données indispensables pour mieux comprendre comment l'enchaînement des différentes acquisitions de l'évolution a ouvert les portes du rêve chez l'homme.
Résumé
Le sommeil humain est le résultat d'une très longue évolution au cours de laquelle les mécanismes suivants ont été intégrés: une horloge circadienne respon sable d'une homéostasie prédictive, des mécanismes d'homéostasie réactive adaptant la durée et/ou l'intensité du sommeil à la durée de l'éveil précédent. Enfin, depuis l'apparition de l'homéothermie, un pace maker ou oscillateur ultradien responsable du sommeil paradoxal est apparu au cours du sommeil. Il est respon sable chez l'homme de l'activité onirique.